Условно разграничивая каскадные ступени Развития Материи,
необходимо чётко себе представлять, что начало этапа фн. развития Материи по каждому
последующему оргуровню и прекращение её развития по предыдущему значительное время
протекают параллельно, одновременно одно с другим. Образование и накопление гумусового
слоя почвы на Земле происходило в течение многих сотен миллионов лет. При этом процесс
протекал одновременно с началом развития биосферы и появлением Жизни на нашей планете.
Формирование биосферы шло главным образом по пути синтеза фщ. единиц гумусового горизонта
почвы, который накапливает и хранит фн. системы - комплексы оргуровня Ж, ставшими
на определённом этапе фщ. единицами уровня З, из которых, в свою очередь,
впоследствии началось образование систем данного подуровня - аминокислот, белков
и других внутриклеточных структур.
Всё это произошло в период, когда, как известно, углеводороды
и их простейшие кислородные и азотистые производные, возникшие на поверхности Земли,
находясь в водном растворе - в первичной земной гидросфере, - в силу действия законов
движения Материи в качестве ![]() )
)
привела к созданию коацерватных капель - индивидуальных белковых комплексов,
отделённых от окружающей среды определённо выраженной поверхностью.
В коацерватных каплях, как и в любой фн. системе Материи
данного организационного уровня, постоянно протекают химические процессы синтеза и
распада. Однако время протекания каждой отдельной реакции под влиянием включённых в
систему катализаторов настолько мало, а частота реакций настолько велика, что процессы
длятся практически беспрерывно. От этого создаётся впечатление "живости"
рассматриваемого объекта. Таким образом, скорости синтеза и распада высокомолекулярных
органических соединений являются основой функционирования всех существующих жизненных
систем, при этом каждая из протекающих реакций имеет свой строго определённый алгоритм.
Соотношение частоты и скоростей указанных процессов зависит от индивидуального состава
и организации каждой данной системы, а также её взаимодействия с условиями окружающей
среды. Если в этом соотношении соблюдается баланс, коацерватная капля, как и любая
система, приобретает динамически устойчивый характер. В случае, если в ней преобладает
скорость и частота синтетических реакций, она растёт. В противом случае она распадается
на составляющие фщ. единицы. Таким образом, существует тесная связь между индивидуальной
системной организацией данной коацерватной капли, теми химическими превращениями, которые
совершаются в ней в соответствии с определёнными для её фн. ячеек алгоритмами, и её
дальнейшей судьбой в данных условиях существования.
В первичной земной гидросфере коацерватные капли,
образовавшиеся путём синтеза белковых молекул, плавали не просто в воде, а в
растворе разнообразных органических и неорганических веществ, то есть готовых
фщ. единиц (уровней Е - Ж). В соответствии с законами движения
Материи в качестве ![]() )
)
Как известно, начало возникновения простейших жизненных систем
произошло около двух млрд. лет назад в протерозойскую эру. Первичные живые существа
зародились в воде в процессе продолжительного развития динамически устойчивых коацерватных
капель, фн. комплексы которых включались в качестве составных частей в системы последующих
оргуровней. Вследствие этого уже на данном этапе Развития Материи наиболее полно проявился
и продолжал своё дальнейшее совершенствование механизм построения высокоорганизованных
систем, один из основных принципов которого заключается в заполнении фн. ячеек системы
не единичными фщ. единицами, а целыми блоками или комплексами их. Под его действием фн.
системы оргуровня З поглощали постоянно окружающие их белковые комплексы,
"расщепляли" их и заполняли образовавшимися блоками свободные фн. ячейки
своих структур, в конечном итоге синтезируя из них фщ. единицы более высокого оргуровня.
При этом энергия, выделяющаяся при расщеплении комплексов, использовалась в большей её
части на осуществление реакций синтеза. Всё это, в конечном итоге, привело к древнейшим
формам организации Жизни, к которым следует отнести бактерии, различные типы водорослей
и грибов. Результатом всего исторического Развития Материи по оргуровню З на
протяжении многомиллионного периода на сегодняшний момент времени являются современные
нам растительные и животные организмы, включая Человека. Мы не будем подробно
рассматривать все этапы филогенеза растительного и животного мира, которые хорошо
известны. Остановимся лишь на основных особенностях движения Материи в качестве
на этих организационных уровнях с тем, чтобы убедиться в том, что и они неразрывно
связаны с закономерностями Развития Материи по всем предыдущим подуровням, являются
их прямым продолжением, неотделимым от них, и вместе с ними составляют единую
развивающуюся системную организацию материальной субстанции.
Итак, Жизнь возникла в результате сложной системной интеграции
фщ. единиц всех подуровней, относящихся к числу так называемых "неорганических"
элементов. Процесс этот протекал направленно в течение длительного периода времени
и состоял, наряду с совершенствованием пространственной структуры фн. ячеек любого
подуровня, в подборе и закреплении оптимального набора алгоритмов для каждой из этих
ячеек, а также оптимального периода функционирования для заполняющих их фщ. единиц.
Деление веществ на неорганические и органические носит довольно условный характер,
но принято считать, что большинство соединений, в состав которых входит углерод,
относятся к разряду органических, так как в природе они встречаются почти исключительно
в организмах животных и растений, принимают участие в жизненных процессах или же являются
продуктами жизнедеятельности или распада организмов.
При всём многообразии природных органических веществ они обычно
состоят из большого числа однотипных элементов - фщ. единиц предыдущих подуровней; в их
состав кроме углерода почти всегда входит водород, часто кислород и азот, иногда сера и
фосфор. Эти элементы называются органогенами, то есть порождающие органические молекулы.
Среди органических соединений широкое распространение получило явление изомерии, то есть
структурное разнообразие системного построения фн. ячеек. В результате при одном и том же
количественном наборе фщ. единиц системы обладают совершенно различными фн. свойствами.
Поэтому явление изомерии, в частности, обусловливает огромное многообразие органических
веществ, вместе с тем ещё более повышая коэффициент полифункциональности фщ. единиц, что
отвечает требованию ускоренного движения Материи в качестве, характерного для данного
оргуровня. Одной из важных особенностей органических соединений, которая накладывает
отпечаток на все их химические свойства, является характер связей между атомами в их
молекулах. В подавляющем большинстве эти связи имеют ярко выраженный ковалентный характер.
Поэтому органические вещества в большинстве неэлектролиты, не диссоциируют в растворах
на ионы и сравнительно медленно взаимодействуют друг с другом. Время, необходимое для
завершения реакций между органическими веществами, обычно измеряется часами, а иногда
и днями. Вот почему в органической химии участие различных катализаторов имеет особенно
большое значение.
Многие из известных органических соединений выполняют функции
носителей, участников или продуктов процессов, протекающих в живых организмах, или же -
такие, как ферменты, гормоны, витамины и др. - являются биологическими катализаторами,
инициаторами и регуляторами этих процессов. Согласно теории химического строения
органического вещества, функциональные свойства соединений зависят от:
1) набора фщ. единиц, определяющего качественный
и количественный их состав;
2) структурного расположения в пространстве
фн. ячеек системы, влияющего на химические свойства вещества;
3) совокупности алгоритмов фн. ячеек данной
системы, которые определяют порядок
а) последовательного
заполнения фн. ячеек соответствующими фщ. единицами,
б) их функционирования и
в) последующего распада
подсистем.
Многообразие органических соединений обусловливается прежде
всего фн. свойствами атомов углерода соединяться прочными ковалентными связями друг
с другом, образуя углеродные цепи практически неограниченной длины.
В процессе Развития Материи по оргуровню З постепенно формировались органические соединения, представлявшие собой всё более динамически устойчивые фн. системы, которые, в свою очередь, затем становились фщ. единицами в системах более высшего порядка. К таким динамически устойчивым органическим соединениям можно отнести, в частности, аминокислоты. Общая формула их построения такова:
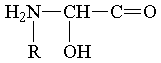
где R - фн. ячейка углеводородного радикала, которую могут занимать и другие различные фщ. единицы.
Из сотен и тысяч молекул аминокислот (как фщ. единиц)
синтезируются более сложные молекулы белковых веществ, или белков (фн. систем),
которые по истечении периода их функционирования под влиянием минеральных кислот,
щёлочей или ферментов распадаются на составляющие их фщ. единицы - аминокислоты
с тем, чтобы дать им затем возможность вновь войти в состав образующихся новых
соединений, то есть заполнить соответствующие им новые фн. ячейки. И процесс этот
повторяется постоянно бесконечное число раз.
О важном значении белков также хорошо известно. Они играют
первостепенную роль во всех жизненных процессах, являются носителями Жизни. Белки в
качестве фщ. единиц сами входят в состав более сложных систем и подсистем организмов,
содержатся во всех клетках, тканях, в крови, в костях и т.д. Ферменты (энзимы), многие
гормоны представляют собой сложные белки.
Всё многообразие белков образовано различными сочетаниями
20 аминокислот; при этом для каждого белка строго специфичной являются структурное
построение системы фн. ячеек, заполняемых соответствующими аминокислотами и другими
фщ. единицами, а также совокупность его алгоритмов, то есть временная последовательность
развёртывания системы белка (заполнение её фн. ячеек фщ. единицами), функционирования и
распада её подсистем. В структуре белковых систем различают подсистемные блок-образования
пептидов, в состав которых входит две или более аминокислот, соединённых пептидными
связями
Дальнейшее совершенствование структур белковых
систем происходило путём объединения полимеров аминокислот в пептидные цепи и
циклические образования в сочетаниях, имеющих различные количественные соотношения
и последовательность фн. ячеек. Образовавшееся в результате этого процесса неисчерпаемое
разнообразие химических структур макрополимеров аминокислот, каждая из которых являлась
сложнейшим системным сочетанием входящих в неё фщ. единиц всех организационных подуровней,
представляло собой в то же время новую группу фщ. единиц более высокого порядка, готовую
заполнить предназначавшиеся для неё соответствующие фн. ячейки новых гиперсистем. При
этом каждая фщ. единица - белок обладала своими, строго индивидуальными особенностями
построения, неизменным количеством фн. ячеек её структуры, строго определённым их
сочетанием и алгоритмами построения, функционирования и распада, что придавало каждой
фщ. единице присущие только ей фн. свойства, соответствующие определённой точке на
координатной прямой движения Материи в качестве.
Одновременно продолжал увеличиваться коэффициент
полифункционирования отдельных фщ. единиц. Принцип действия механизма
полифункционирования сводится к следующему. Если взять какую-либо фщ. единицу
с определёнными фн. свойствами и помещать её последовательно то в одну, то в
другую фн. ячейку, и она при этом сможет нормально выполнять необходимые для
данных фн. ячеек алгоритмы, то это и будет означать, что ей присуще свойство
полифункционирования. Чем большее количество фн. ячеек различных структур может
попеременно занимать данная фщ. единица в некоторый промежуток времени, тем выше
её коэффициент полифункционирования. Как правило, каждая единица единовременно может
занять только одну фн. ячейку какой-либо структуры. В качестве примера можно назвать
любой химический элемент типа водорода, кислорода, хлора, которые могут входить в
состав многих химических соединений, но в данный конкретный момент пребывают только
в одном из них. Другой разновидностью полифункционирования является изъятие из
какой-либо фн. ячейки системы фщ. единицы x и помещение туда фщ. единицы
y или z, вследствие чего фн. свойства данного системного
образования соответственно изменятся. При обратном перемещении фщ. единиц система вновь
обретает свои первоначальные фн. свойства; и поэтому, чем большую замену фщ. единиц в
своих ячейках допускает в некоторый отрезок времени ![]() данная система, тем выше коэффициент ее
полифункционирования. В этом случае примерами могут служить все обратимые химические
реакции замещения типа
данная система, тем выше коэффициент ее
полифункционирования. В этом случае примерами могут служить все обратимые химические
реакции замещения типа
Аминокислоты, входя в состав белковой молекулы, сохраняют
свободными и реакционноспособными свои специфические полифункциональные ячейки,
химические функции которых состоят в способности присоединять различные системные
группировки. Это обусловливает взаимодействие белков с самыми различными веществами,
создаёт исключительные химические возможности, которыми не обладают никакие другие
вещества данного подуровня. В силу этого белки, входящие, например, в состав живой
протоплазмы, сочетаются в комплексы с другими соединениями - от воды и минеральных
веществ до всевозможных органических соединений, в том числе и с другими белками. Эти
комплексы, в зависимости от образующего их фактора, могут быть довольно устойчивыми
и образовываться в количествах, необходимых для построения гиперсистем. Примерами
таких комплексов служат разнообразные сложные белки - нуклеопротеиды, хромопротеиды,
липопротеиды, металлопротеиды и т.п. Они участвуют в построении гиперсистемных структур
и, вместе с тем, играют большую роль в их функционировании благодаря своим каталитическим
свойствам. Наряду с устойчивыми соединениями белки способны образовывать и крайне
эфемерные комплексы, период функционирования которых сравнительно небольшой. Подчиняясь
соответствующим алгоритмам, эти соединения быстро возникают и, отфункционировав,
также быстро разлагаются. Таким образом, через механизм полифункциониравания самые
разнообразные элементы из аккумулятивных подсистем вовлекаются в обмен веществ живой
организации Материи для временного использования их фн. свойств в том или ином
системном образовании.
При заполнении фн. ячеек многомолекулярных соединений
отдельными индивидуальными белками - фщ. единицами образуются новые системные единицы,
физические и химические свойства которых существенно отличаются от свойств входящих в
их состав отдельных белков. Ассоциируясь между собой, белки образуют целые молекулярные
рои, представляющие собой различные структурные образования живого вещества. Весьма
существенным является и то, что фн. свойства белков, их способность реагировать с
разнообразными веществами и ассоциироваться в многомолекулярные комплексы определяется
не только составом и расположением аминокислотных остатков, но и пространственной
конфигурацией белковой молекулы, то есть относительным расположением в пространстве
отдельных частей её структуры. Химическое взаимодействие боковых радикалов и полярных
групп аминокислотных остатков, действуя внутримолекулярно, приводит к закономерному
скручиванию пептидных цепей белковой молекулы и объединению их в клубки, в так
называемые белковые глобулы, обладающие упорядоченной пространственной конфигурацией.
Во внутреннем строении белковых глобул отдельные участки пептидных цепей и замкнутых
колец оказываются определённым образом расположенными по отношению друг к другу и
взаимно закреплёнными путём сшивания этих участков водородными или другими прочными
связями. Такого рода строение обусловливает определённые размеры и форму белковых
глобул. Она может приближаться к шаровидной или быть сильно вытянутой. Те или иные
изменения окружающей глобулу внешней среды сильно влияют на её форму, сильно сжимая
или, наоборот, растягивая её. В зависимости от того, какие активные группировки фщ.
единиц аминокислотных остатков при данной конфигурации глобулярного клубка оказываются
расположенными на поверхности и, следовательно, доступными химическому взаимодействию
и какие будут скрыты в глубине, защищены, "экранированы" соседними
группировками, зависят изменяющиеся фн. свойства белка, даже при сохранении постоянным
его аминокислотного состава. Поэтому даже очень небольшие изменения пространственной
архитектоники глобулы оказывают решающее влияние на химическую реактивность белка
и на те тонко нюансированные его свойства, которые определяют собой биологическую
специфичность каждого индивидуального белкового соединения. Этот созданный в процессе
Развития Материи ещё один, более сложный и тонкий механизм полифункционирования
способствовал диктуемому законами Развития ускоренному движению Материи по категории
качества ![]() ).
).
Известно, что химические реакции между органическими
соединениями совершаются в живых организмах с очень большими скоростями, хотя и вполне
измеримыми, но совершенно несравнимыми с теми, которые наблюдаются при взаимодействии
этих соединений в изолированном и очищенном виде вне структур живых тел. Причина
этого заключается в том, что в составе живой протоплазмы всегда присутствуют особые
биологические ускорители - ферменты, называемые протеинами (простые белки) или
протеидами (сложные белки), в которых белок соединен в комплексе с небелковой
("простетической") группой - в большинстве случаев с металлоорганическим
соединением или тем или иным витамином. В силу этого в каждой живой клетке присутствует
целый набор разнообразных ферментов, поскольку ферментативной активностью обладает
большинство протеинов и протеидов живого организма. Таким образом, ферменты составляют
основную массу протоплазменных белков. То обстоятельство, что основой ферментных
комплексов всегда являются обладающие определённой архитектоникой те или иные
белковые глобулы, обусловливает ряд особенностей, которые отличают ферменты от других
катализаторов. Это прежде всего исключительная каталитическая мощь ферментов. Известно
большое число неорганических и органических соединений низших организационых уровней,
способных ускорять те же реакции, что и ферменты. Механизм действия любого катализатора
весьма прост и напоминает действие ключа, вводимого в ту или иную систему. При реакциях
распада свободные связи катализатора нейтрализуют силы связи, объединяющие вместе фщ.
единицы в единую систему, и она распадается на составные части. В реакциях синтеза
катализатор путём предоставления своих свободных связей ускоряет процесс объединения
фщ. единиц. Однако, сложность и совершенство системной структуры ферментов намного
повысили силу их каталитического воздействия по сравнению с менее организованными
катализаторами, что нашло своё отражение в сокращении времени протекания реакций, то
есть перестроения структуры-принципала. Так, например, ион железа разлагает перекись
водорода на кислород и воду. Соответствующий фермент (каталаза), представляющий собой
сочетание железо-порфиринового комплекса со специфическим белком, обладает тем
же действием. Но он осуществляет эту реакцию в десять миллиардов раз скорее, чем
неорганическое железо. Иными словами, 1 мг железа, включённый в ферментный комплекс,
может по своему каталитическому действию заменить 10 тонн неорганического железа.
Таким образом, ферменты являются относительно сложными системными образованиями
уровня З, функция которых заключается в обеспечении регулирования в определённом
диапазоне времени структурных перестроений гиперсистем, в которые они входят, в
соответствии с предписаниями усложняющихся алгоритмов гиперполифункционирования, то
есть корреляции системных структур в зависимости от изменения их фн. свойств. Поэтому
даже незначительные изменения в структурном строении ферментного комплекса, перестановка
тех или иных радикалов в простетической группе или нарушение архитектоники белкового
компонента приводят к резкому понижению каталитической активности данного фермента.
Следовательно, в системной организации ферментов также подтверждается то соответствие
между структурным построением фн. ячеек и функцией всей данной системы, которое является
закономерным для всех ступеней и уровней каскадного Развития Материи вообще.
Пространственная конфигурация белковых глобул определяет
собой и вторую особенность ферментов - высокую специфичность их действия, то есть
монофункционирование. Иными словами, каждый фермент способен катализировать только
свою, строго определённую реакцию. Поэтому, если имеется какое-либо органическое
вещество, способное к ряду химических соединений, то в присутствии того или иного
фермента оно будет быстро реагировать только в одном, строго определённом направлении,
выполняя тем самым соответствующий алгоритм данной системы.
Наконец, специфическое строение белков обусловливает собой
и третье характерное для ферментов свойство - их исключительную чувствительность
к различного рода воздействиям. Так, при определённых физических или химических
воздействиях самого различного рода (даже тогда, когда эти воздействия не затрагивают
пептидных и других ковалентных связей белковой молекулы) специфическая пространственная
архитектоника глобулы может измениться и даже нарушиться, а её упорядоченная структурная
конфигурация необратимо утратится. В этом случае пептидные цепи принимают беспорядочное
пространственное расположение и белок из глобулярного состояния переходит в фибриллярное
- происходит так называемая денатурация белков, при которой они теряют ряд тех своих
специфических биологически важных свойств, которые обусловливаются определённой
архитектоникой каждого типа белковой молекулы. Совершенно исчезают при этом
ферментативные свойства белков. Однако при более мягких воздействиях каталитическая
активность ферментного комплекса может до известной степени сохраняться, претерпевая
лишь те или иные количественные сдвиги. Поэтому любые, даже весьма незначительные,
изменения физических или химических условий в той среде, где протекает данная
ферментативная реакция, всегда находят своё отражение в изменении её характера
и скорости. Все эти свойства белков составляли основу качественного Развития Материи
по организационному уровню З, в системах которого происходила всё большая фн.
дифференциация фщ. единиц и структурная интеграция фн. ячеек.
Каждая фщ. единица, попав в соответствующую ей фн. ячейку,
функционирует в ней определённый алгоритмами период, после чего покидает её, уступая
место новой фщ. единице с теми же фн. свойствами. Покинув одну фн. ячейку, фщ. единица
перемещается в предписанную ей алгоритмами другую ячейку и т.д. Процесс этот происходит
постоянно, периодически возобновляясь и повторяясь, отчего создаётся впечатление движения
фщ. единиц - веществ через системную структуру каждого данного образования, во время
которого система поглощает фщ. единицы (или их комплексы), некоторое время использует
их внутри себя, а затем выводит за свои пределы. Это непрекращающееся движение
регулируется и регламентируется совокупностью соответствующих алгоритмов каждой
системы, в то время как постоянно протекающие в системе реакции придают ей своеобразную
"живость". В силу этого при так называемом обмене веществ очень простые
и порой однообразные химические реакции окисления, восстановления, гидролиза,
фосфоролиза, альдольного уплотнения, разрыва углеродной связи и т.д. (которые могут
быть воспроизведены и вне системы организма) определённым образом организованы и
сочетаются во времени соответствующими алгоритмами, а также подчинены функциональным
интересам своей системы, как интегрированного единого целого. Эти реакции протекают
в системах уровня З не случайно, не хаотически, а в строго определённой
взаимопоследовательности, зафиксированной алгоритмами. То колоссальное разнообразие
органических соединений, которое к настоящему времени представлено в мире живых
существ, обусловлено не разнообразием и сложностью отдельных индивидуальных реакций,
а разнообразием их сочетаний, изменением той последовательности, в которой они протекают
в любой клетке живого организма на той или иной стадии его развития. Другими словами,
развитие систем данного уровня организации Материи оказалось в ещё большей зависимости
от появления новых алгоритмов, совершенствования структур фн. ячеек и своевременного
заполнения их соответствующими фщ. единицами. Последовательность химических реакций,
обусловленная соответствующими алгоритмами, лежит в основе как синтеза, так и распада
живого вещества, в основе таких жизненных явлений, как брожение, дыхание, фотосинтез
и т.д. Молекулы сахара и кислорода, углекислоты и воды являются в этом случае лишь
начальными и конечными звеньями в длинной цепи химических превращений, причём возникающее
в результате одной реакции промежуточное вещество (фщ. комплекс) сейчас же вступает в
следующую строго определённую для данного жизненного процесса реакцию. При изменении
этой последовательности, при исключении или замене хотя бы одного какого-нибудь звена
в цепи превращений, предопределённых данным алгоритмом, весь процесс в целом совершенно
изменяется или даже полностью нарушается.
В основе механизма этих явлений лежит тесная согласованность
скоростей отдельных химических реакций, представляющих собой перемещение фщ. единиц
нижних подуровней из одних фн. ячеек в другие. Любое органическое вещество может
реагировать в очень многих направлениях, то есть обладает весьма большими и
разнообразными возможностями, однако их реализация может осуществляться с очень
различными скоростями, в зависимости от всей совокупности тех условий, в которых
данная реакция протекает. Понятно, что если в данных условиях одна какая-либо реакция
осуществляется весьма быстро, а все остальные потенциально возможные реакции происходят
относительно медленно, то практическое значение этих последних реакций в общем процессе
обмена оказывается совершенно ничтожным. Иными словами, перед каждым органическим
веществом протоплазмы открыты многочисленные пути химических превращений, но фактически
в обмене веществ каждое поступающее из внешней среды соединение или каждый образующийся
промежуточный продукт будут изменяться только в том направлении, в котором они реагируют
с наибольшей скоростью. Все остальные, медленно протекающие реакции просто не успевают
за то же время реализоваться в сколько-нибудь значительных размерах.
Вступающие в процесс обмена веществ в качестве реагентов фщ.
единицы - субстрат заполняют собой строго предназначенные для них фн. ячейки в структуре
данной системы, в которых в определённый момент времени по предписанию алгоритмов они
вступают в комплексное соединение с соответствующим белком-ферментом. Каждый такой
комплекс является неустойчивым образованием, но достаточно надёжным, чтобы выполнить
какую-нибудь необходимую функцию. Отфункционировав, он очень быстро подвергается
дальнейшему превращению, при этом субстрат изменяется в соответствующем направлении,
то есть составляющие его фщ. единицы переходят в другие фн. ячейки, а фермент
регенерирует и может опять вступить в комплекс с новой порцией субстрата для поддержания
возможности выполнения необходимой функции данным системообразованием. Поэтому для того,
чтобы любая фщ. единица реально могла участвовать в обмене веществ в системах уровня
З, она должна войти во взаимодействие с белком, образовать с ним определённое
комплексное соединение и только таким путём реализовать свои фн. свойства. В силу этого
то направление, в котором изменяется при обмене веществ любое органическое соединение,
зависит не только от индивидуального молекулярного строения составляющих фщ. единиц и
определяющих его фн. свойства, но и от фн. ячейки, куда каждая фщ. единица соединения
попадает и где она совместно с другими фщ. единицами - белками должна образовать фн.
комплекс с новыми фн. свойствами, способный выполнить ту или иную новую функцию,
подчиняясь превалирующим в данной системе алгоритмам.
Вследствие чрезвычайно тонкой специфичности ферментных белков,
каждый из них, обладая строго индивидуальными фн. свойствами, может попасть только в
строго определённые фн. ячейки и, в силу этого, способен образовывать фн. комплексы
только с определёнными фщ. единицами предыдущих подуровней, а также катализировать лишь
определённые индивидуальные реакции. Поэтому в осуществлении того или иного жизненного
процесса, а тем более всего обмена веществ в целом, участвуют тысячи индивидуальных
белков - ферментов, при этом каждый из них способен специфически катализировать лишь
отдельную реакцию, и только в совокупности, в определённом сочетании своего действия
они создают тот закономерный порядок явлений, который лежит в основе процесса обмена
веществ. Итак, обмен веществ, происходящий постоянно в системах любого жизненного
организма, это сложнейший клубок химических превращений обмена, где регламентируемые
данной совокупностью алгоритмов сплетаются в едино действующий механизм тысячи
индивидуальных реакций, суть каждой из которых сводится к перемещению той или
иной фщ. единицы из одной фн. ячейки структуры системы в другую, при этом моменты
перемещения фщ. единиц по ячейкам строго согласованы по всей системе, чередуются в
строго определённом порядке и в каждом перемещении участвуют строго означенные фщ.
единицы и фн. ячейки. Вместе с тем, большую роль для течения каждой реакции обмена
веществ играет внесистемная и околоподсистемная среда или, иными словами, системное
окружение единицами предыдущих подуровней Материи. Так, всякое повышение или понижение
температуры, всякое изменение кислотной среды, окислительного потенциала или
осмотического давления смещает соотношение между скоростями отдельных ферментативных
реакций, происходящих в системе данного живого организма, а, следовательно, изменяет
их взаимосвязь во времени, что, в свою очередь, находит отражение в изменении периодов
функционирования тех или иных фщ. единиц. Таким образом, системная организация живого
вещества неразрывно связана с околосистемной организацией среды и составляет с ней
единое целое. Кроме того, очень большое влияние на порядок и направление лежащих в
основе обмена ферментативных реакций имеет и пространственная организация фн. ячеек
в структуре живого вещества. Итак, многие десятки и сотни тысяч химических реакций,
непрерывно протекающих в каждом живом организме, не только строго согласованы между
собой во времени бесчисленное число раз отработанными алгоритмами, не только
сочетаются в едином порядке всей структурной организацией его системы и окружающей
его околосистемной среды, но и сам весь этот порядок направлен на поддержание в
течение определённого периода времени гиперфункциональных свойств всей данной
системы в целом, как фщ. единицы более высокого уровня. Вновь приобретённые при
этом фн. свойства белковых веществ могут стать ясными лишь при изучении особенностей
их функционирования в организме в качестве фщ. единиц систем более высокого
организационного уровня Материи.
В связи с тем, что с момента вступления качественных форм Материи
в так называемую "живую" стадию Развития характер организации систем усложнился,
помимо организующих начал, характерных для систем предыдущих подуровней, как то:
1) наличие строго регламентированного количества
фн. ячеек, объединённых в единую структуру связей,
2) заполняющих их и соответствующих им фщ. единиц,
3) совокупности алгоритмов
построения, функционирования и распада,
4) энергообеспечения процесса функционирования системы
для организационного уровня З потребовались дополнительные системообразующие
факторы. Ввиду большей усложнённости его фн. систем происходило увеличение их кажущейся
автономности, которая фактически представляет собой лишь больший разрыв в уровнях
организации самой системы и околосистемной среды и которая дала повод обозначать
их некоторые свойства с приложением полуслова "само": самообновление,
саморегулирование, самоэнергообеспечение и чуть ли ни самоуничтожение. Основами этой
автономности явилось начало развития соответствующих подсистем в общей структуре
организма, отвечающих за обеспечение той или иной специфической функции. Происходившее
в силу дальнейшей дифференциации функций всё большее расслоение систем на подсистемы
ещё более усложнило структуру систем и потребовало более чёткой взаимокоординации её
интегрированных составных частей. Поэтому совокупность алгоритмов каждой системы
постепенно увеличивалась в количественном выражении, ещё более улучшался её качественный
состав.
Всем известно, что такое алгоритм. Это строго
регламентированный во времени и в пространстве порядок последовательного перемещения
фщ. единиц из одной фн. ячейки структуры данного уровня в другую. Этот порядок обязателен
для систем любого организационного уровня, предопределён для каждой их фщ. единицы. Всё
вокруг нас подчинено тем или иным алгоритмам. Их великое множество - от самых простых
до невероятно сложных. Среди простых бытовых алгоритмов мы можем назвать алгоритмы
приготовления пищи (например, заварки чая, выпечки пирогов и т. п.), изготовления стола
или скамейки, выращивания картофеля и т. д. Среди суперсложных можно назвать, например,
алгоритм изготовления авианосца. Поэтому в обычной поварской книге перечислены алгоритмы
приготовления пищи, в нотах - алгоритмы воспроизведения музыкальных произведений, а
в технологических картах построения жилого дома или автомобиля, прокладки дороги -
алгоритмы их построения. Все указанные нами алгоритмы были выработаны в течение
практической деятельности человеком. Однако, кто же занимался составлением алгоритмов
для построения фн. систем доорганической и органической организации Материи? Ведь
уже алгоритмы построения атома водорода или молекулы аминокислоты являются довольно
непростыми. Конечно, их никто не изобретал. Они вырабатывались сами, повинуясь железной
необходимости, вытекающей из действия законов Развития Материи, и в первую очередь, её
движения по категории качества ![]() ).
).
По мере усложнения системных структур уже в начальный период
организации живых форм Материи, продолжительность функционирования которых основана,
как известно, на принципе постоянной замены в них блоков фщ. единиц, в некоторый
момент организационного развития потребовался механизм, обеспечивающий создание таких
блоков в сравнительно короткое время с тем, чтобы заменять ими отфункционировавшие в
фн. ячейках блоки без нарушения фн. свойств всей данной системы в целом. С этой целью
в системах стала всё более выделяться подсистема, записывающая алгоритмы построения
того или иного блока, их пространственного расположения в общей структуре и временной
последовательности перехода фщ. единиц данного уровня из одних фн. ячеек в другие. Как
известно, в доорганических системах их структуры имели долговременный характер, при
этом эти суммативные системные образования составлялись из фщ. единиц нижних подуровней
в соответствии с их, главным образом, физическими свойствами при одновременном
аккумулировании большого количества энергии. Распад таких систем происходил через
большой отрезок времени, имел разовый нерегулярный характер и служил лишь целям общего
перестроения макросистемы в целом. Позднее, на молекулярном организационном уровне
порядок составления системных образований помимо физических стал регулироваться также
и химическими свойствами входящих в них фщ. единиц, при этом с повышением системной
организации происходило всё меньшее аккумулирование суммарной энергии (хотя из расчёта
на одну фщ. единицу каждого последующего уровня аккумулирование энергии значительно
возрастало), а сами соединения носили всё более кратковременный характер. В
надмолекулярных системах, обладавших всё большим количеством органических свойств,
запись информации об алгоритмах построения и функционирования стали принимать
на себя фн. подсистемы, условно названные впоследствии нуклеотидами.
Итак, в процессе Развития Материи по организационному уровню
З на отдельных участках поверхности планеты Земля с определённого момента времени
стали появляться высокомолекулярные материальные образования, способные нести различную
функциональную нагрузку нового спектра. Они включали в структуры своих подсистем следующие
органические химические соединения: белки, жиры, углеводы, нуклеиновые кислоты и другие
низкомолекулярные органические вещества. Кроме того, в них входили и неорганические
вещества, главным из которых была вода. По мере продвижения актуальной точки Развития
Материи по ординате времени, число новых системных образований сбалансированно
увеличивалось, совершенствовалась их системная структура. Системы уровня З
не были организационно оторваны от предыдущих уровней, а органически включали их
системные образования в качестве фщ. единиц в свои фн. ячейки. Ввиду того, что
пространственное развитие систем оргуровня З было ограничено не только площадью
Земной поверхности, но также и другими факторами физического и химического характера
(такими, как уровень получаемой лучистой энергии Солнца, различный на разных участках
Земной поверхности; наличие в данном месте необходимого спектра системных образований
предыдущих уровней и т.д.), постоянно существовало положение, при котором
![]() . Вследствие этого
Развитие Материи вынуждено было осуществляться практически только за счёт движения по
координате качества
. Вследствие этого
Развитие Материи вынуждено было осуществляться практически только за счёт движения по
координате качества ![]() ),
),
Как известно, у разных клеток обнаруживается сходство не
только в строении, но и в химическом составе, что указывает на то, что их происхождение
было подчинено единым законам Развития Материи. Среднее содержание химических элементов
в клетках таково (в %):
|
|
Из 104 элементов периодической системы Менделеева в клетках обнаружено более 60. Атомы кислорода, углерода, водорода и азота заполняют 98% фн. ячеек клеточных подсистем. 1,9% предоставлены атомам калия, серы, фосфора, хлора, магния, натрия, кальция и железа. Менее 0,1% фн. ячеек занято прочими веществами (микроэлементами). Различные сочетания указанных элементов дают несколько типов внутриклеточных подсистемных образований, которые каждая клетка включает в свои фн. ячейки в качестве фщ. единиц в следующих пропорциях (в %):
|
|
|||||||||||||||||||
Все указанные вещества, сами сложные в структурном
отношении, не нагромождены в клетке вместе в хаотическом беспорядке, а в качестве
фщ. единиц заполняют расположенные в строго определённом порядке предназначенные
для каждого из них фн. ячейки её единой структуры. Функционируя, они проделывают
свои чётко определённые микродвижения внутри микрообъёма пространства клетки,
регулируемые соответствующими внутриклеточными алгоритмами, при этом существует
безусловная связь этих движений в пространстве как с абсолютным, так и с относительным
течением времени. Каждое из веществ клетки в качестве фщ. единицы несёт строго
определённую функцональную нагрузку и имеет свои, регламентируемые соответствующими
алгоритмами, периоды функционирования. Всё их разнообразное сочетание представляет
собой единый тонко отрегулированный клеточный механизм.
К наиболее простым структурным внутриклеточным образованиям
относятся углеводы, жиры, липоиды. Фн. ячейки их структур заполняют, в основном, атомы
углерода, водорода и кислорода. Функция углеводов наиболее проста. Распадаясь
на CO2 и воду с выделением из 1 грамма 4,2 ккал энергии, они обеспечивают
основной массой этих фщ. единиц соответствующие фн. ячейки структуры клеток.
Роль жировых соединений более сложна. Они придают
клеткам гидрофобные (водоотталкивающие) свойства, являются теплоизоляторами. В случае
необходимости, они, как и углеводы, являются источником аккумулированной энергии,
расщепляясь до CO2 и H2O. Расщепление 1 грамма жира даёт 9,3 ккал.
Ещё более сложными структурными образованиями являются
белки. Помимо углерода, водорода и кислорода в фн. ячейках их структур имеются
также атомы азота, серы и других веществ. Белки являются макромолекулами, объединяющими
десятки, сотни тысяч атомов. (Так, если молекулярная масса бензола равна 78, то белка
яйца - 36.000, белка мышц - 1.500.000 и т.д.).
Системная организация белков имеет свои особенности. Входящие
в них атомы заполняют предназначенные для них фн. ячейки не по одному, а целыми
аминокислотными блоками, имеющими устойчивый характер внутрисистемной связи. Всего таких
фщ. единиц - блоков 20. Все они имеют различную системную структуру и выполняют различные
функции. Поэтому образование белков носит поэтапный характер. Вначале образуются
аминокислоты, которые посредством пептидных связей соединяются в белковые цепи с
выделением воды. В среднем каждая белковая цепь содержит до 200 - 300 аминокислотных
блоков в различных сочетаниях. Достаточно в цепи заменить один тип аминокислоты
на другой, как вся структура данного белка, а с ней и его функциональные свойства,
меняются. Структура белковой цепи аминокислотных блоков имеет форму глобул, что придаёт
длинным цепям белка компактный вид и мобильность при пространственном перемещении. В
укладке полипептидной цепи нет ничего случайного или хаотичного, каждому белку присущ
определённый, всегда постоянный характер укладки. Иными словами, структура каждого белка
имеет строго определённое пространственное расположение её фн. ячеек, которые заполняются
строго соответствующими им фщ. единицами - аминокислотными блоками. При этом каждая
структура белка, будучи фщ. единицей в системе более высокого порядка и занимая в ней
соответствующую ей фн. ячейку, выполняет там свою, присущую только ей, функцию. Как
правило, белковые структуры являются активнейшими реагентами химических реакций,
постоянно протекающих внутри клетки, и поэтому наиболее велика их роль в качестве
катализаторов этих реакций. Почти каждая химическая реакция в клетке катализируется
своим особым белком - ферментом, каталитическая активность которого определяется
небольшим участком - его активным центром (сочетанием аминокислотных радикалов).
Структура активного центра фермента и структура субстрата точно соответствуют один
другому. Они подходят друг к другу как ключ к замку. Благодаря наличию структурного
соответствия между активным центром фермента и субстратом они могут тесно сблизиться
между собой, что и обеспечивает возможность реакции между ними.
К другим важным внутриклеточным образованиям следует отнести
нуклеиновые кислоты: дезоксирибонуклеиновую - ДНК и рибонуклеиновую - РНК. Их
основная функция состоит в обеспечении процесса синтеза белков клетки. Длина молекулы
ДНК в сотни и тысячи раз больше самой крупной белковой молекулы и может достигать
нескольких десятков и сотен микрометров, в то время как длина самой крупной белковой
молекулы не превышает 0,1 мкм. Ширина двойной спирали ДНК всего 20
![]() .
Молекулярная масса составляет десятки и даже сотни миллионов. Каждая цепь ДНК - полимер,
мономерами которого являются молекулы четырёх типов нуклеотидов. Иными словами, ДНК - это
полинуклеотид, в цепи которого в строго определённом и для каждой ДНК всегда постоянном
порядке следуют нуклеотиды, являющиеся таким образом фщ. единицами в структуре фн. ячеек
ДНК. Поэтому, если хотя бы в одной из фн. ячеек будет помещена иная фщ. единица -
нуклеотид, фн. свойства всей структуры изменятся. В каждой цепи ДНК (средний мол. вес
10 млн.) содержится до 30 тыс. нуклеотидов (мол. вес 345), вследствие чего количество
изомеров (при 4 типах нуклеотидов) очень велико.
.
Молекулярная масса составляет десятки и даже сотни миллионов. Каждая цепь ДНК - полимер,
мономерами которого являются молекулы четырёх типов нуклеотидов. Иными словами, ДНК - это
полинуклеотид, в цепи которого в строго определённом и для каждой ДНК всегда постоянном
порядке следуют нуклеотиды, являющиеся таким образом фщ. единицами в структуре фн. ячеек
ДНК. Поэтому, если хотя бы в одной из фн. ячеек будет помещена иная фщ. единица -
нуклеотид, фн. свойства всей структуры изменятся. В каждой цепи ДНК (средний мол. вес
10 млн.) содержится до 30 тыс. нуклеотидов (мол. вес 345), вследствие чего количество
изомеров (при 4 типах нуклеотидов) очень велико.
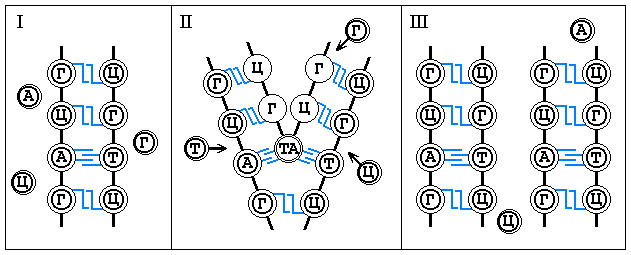
Благодаря принципу комплементарности, лежащему в основе
построения двойной спирали ДНК, молекула ДНК способна к удвоению. Во время этого
процесса две цепи разъединяются, образуя при этом две двойные цепи фн. ячеек, у которых
лишь один ряд заполнен фщ. единицами, а другой становится свободным. На следующем этапе
неассоциированные нуклеотиды из околосистемной среды заполняют соответствующие им
свободные фн. ячейки в обеих спиралях. В результате редупликации вместо одной молекулы
ДНК возникает две такого же точно нуклеотидного состава, как и первоначальная. Одна цепь
в каждой вновь образовавшейся молекуле ДНК остаётся от первоначальной молекулы, другая
синтезируется вновь. Таким путём вместе со структурой происходит передача фн. свойств
ДНК от материнской клетки к дочерней.
Графически это выглядит следующим образом:
Молекулы РНК также являются полимерами, как и ДНК,
но в отличие от них имеют одну спираль фн. ячеек, а не две. РНК выполняет в клетке
ряд функций, в том числе
1) транспортную (транспортируют
аминокислотные блоки к месту синтеза белков);
2) информационную (переносят
информацию о структуре белка);
3) рибосомную.
Ещё одним очень важным нуклеотидом в структуре живой клетки
является аденозинтрифосфорная кислота - АТФ, содержание которой в клетках колеблется
от 0,04 до 0,2 - 0,5%. Его особенность состоит в том, что при отщеплении одной молекулы
фосфорной кислоты АТФ переходит в АДФ (аденозиндифосфорную кислоту) с выделением 40 кдж
энергии с 1 гр-молекулы.
Все указанные выше органические вещества являются сложными по
своей структуре и системной организации образованиями, но и они, в свою очередь, входят
в качестве фщ. единиц в фн. подсистемы интегрированной системы клетки. К числу основных
подсистем клетки отосятся следующие:
Наружная мембрана клетки. Регулирует поступление
ионов и молекул в структуру клетки и выход их из неё в околосистемную среду. Такой обмен
молекулами и ионами, то есть различными фщ. единицами, между системой клетки и внешней
средой происходит постоянно. Различают фагоцитоз - поглощение мембраной крупных частичек
вещества и пиноцитоз - поглощение воды и водного раствора. Через наружную мембрану из
клетки выводятся продукты её жизнедеятельности, то есть отфункционировавшие в подсистемах
клетки фщ. единицы.
Цитоплазма. Внутренняя полужидкая среда клетки,
в системном объёме которой развёрнута внутренняя структура клетки, то есть её ядро, все
органоиды (или органеллы), включения и вакуоли. Цитоплазма состоит из воды с растворёнными
солями и разнообразными органическими веществами, среди которых преобладают белки.
Структура цитоплазмы состоит из фн. ячеек, не связанных жестко и свободно перемещающихся
во всём её объёме. Заполняющие их фщ. единицы, когда это необходимо, переходят из них в
фн. ячейки органоидов. Поэтому основными функциями цитоплазмы являются аккумулятивная и
транспортная.
Эндоплазматическая сеть. Органоид клетки,
представляющий собой сложную систему каналов и полостей, пронизывающих всю цитоплазму
клетки. На мембранах гладкой эндоплазматической сети происходит синтез жиров и углеводов,
которые накапливаются в аккумулятивных фн. ячейках её каналов и полостей, а затем
транспортируются к различным органоидам клетки, где они в качестве фщ. единиц занимают
соответствующие фн. ячейки их структур. На мембранах каналов и полостей располагается
также множество мелких округлых телец-рибосом.
Каждая рибосома состоит из двух мелких частиц, в состав которых
входят белки и РНК. В каждой клетке содержится по нескольку тысяч рибосом. На рибосомах
синтезируются все белки, входящие в состав данной клетки, путём сборки белковых молекул
из аминокислот, имеющихся в цитоплазме. Синтез белков - это сложный процесс заполнения
аминокислотными блоками соответствующих фн. ячеек их структур, который осуществляется
одновременно группой из нескольких десятков рибосом, или полирибосомой. Синтезированные
белки сначала накапливаются в каналах и полостях гранулярной эндоплазматической сети, а
затем транспортируются к тем подсистемам клетки, где расположены предназначенные для них
фн. ячейки. Эндоплазматическая сеть и полирибосомы представляют собой единый механизм
биосинтеза, аккумулирования и транспортировки белков.
Митохондрии. Органоид, основная функция
которого состоит в синтезе АТФ, представляющей универсальный источник энергии, необходимой
для осуществления постянно протекающих внутри клетки химических процессов. Количество
митохондрий в клетке колеблется от нескольких до сотен тысяч. Внутри митохондрий находятся
рибосомы и нуклеиновые кислоты, а также большое количество разнообразных ферментов.
Синтезированная АТФ заполняет транспортные фн. ячейки цитоплазмы и направляется к ядру
и органоидам клетки.
Пластиды. Органоиды растительных клеток.
Бывают различных типов. С помощью одного из них - хлоропластов, благодаря входящему
в их состав пигменту (хролофиллу), клетки растений способны, используя световую энергию
Солнца, синтезировать из неорганических веществ органические (углеводы). Этот процесс,
как известно, носит название фотосинтеза.
Комплекс Гольджи. Органоид всех растительных
и животных клеток, в котором происходит накопление белков, жиров и углеводов с последующей
их транспортировкой в соответствующие фн. ячейки как внутри клетки, так и вне её.
Лизосомы. Органоид, имеющийся во всех клетках
и состоящий из комплекса ферментов, способных расщеплять белки, жиры и углеводы. В этом
заключается основная функция лизосом. В каждой клетке сосредоточены десятки лизосом,
участвующих в расщеплении уже отфункционировавших или аккумулятивных системных
образований, а также тех, которые попадают в клетку извне путём фагоцитоза и пиноцитоза.
В результате расщепления фщ. единицы покидают фн. ячейки расщепляемых структур, собираются
в фн. ячейках аккумулятивных систем данной клетки, а затем транспортируются в фн. ячейки
её новых системных образований. Расщеплённые с помощью лизосом отфункционировавшие
структуры клетки удаляются за её пределы. Образование новых лизосом происходит в клетке
постоянно. Ферменты, функционирующие в лизосомах, как и всякие другие белки, синтезируются
на рибосомах цитоплазмы. Затем эти ферменты поступают по каналам эндоплазматической сети
к комплексу Гольджи, в полостях и трубочках которого формируются фн. ячейки структур
лизосом. Сформировавшись, лизосомы отделяются от концов трубочек и поступают в цитоплазму.
Клеточный центр. Органоид, расположенный в
одном из участков уплотнённой цитоплазмы. В него входят две центриоли, играющие важную
роль при делении клетки.
Структура клеток содержит и другие органоиды: жгутики,
реснички и т.п., а также клеточные включения (углеводы, жиры, белки).
Вместе с тем, клетки, будучи сами по себе очень сложными
системными образованиями, в свою очередь являются фщ. единицами, заполняющими фн.
ячейки гиперсистем последующих уровней организации Материи. Вследствие этого в
системной организации клеток предусмотрен механизм, позволяющий за сравнительно
короткий период времени создавать аналогичные им системные формирования. В результате
клеточный цикл включает два периода:
1) Деление (митоз), в процессе которого
образуются две дочерние клетки;
2) Период между двумя делениями - интерфаза -
собственно время функционирования клетки.
Большую роль в делении клетки играет её ядро, имеющееся
в каждой клетке и представляющее собой сложную фн. подсистему. Ядро имеет ядерную
оболочку, через которую в него и из него поступают белки, углеводы, жиры, нуклеиновые
кислоты, вода и разнообразные ионы. Попав внутрь ядра, они заполяют фн. ячейки ядерного
сока, а также ядрышек и хроматина. В ядрышках происходит синтез РНК, сами же они
формируются только в интерфазе. Хроматин представляет собой однородное вещество,
служащее аккумулятивной подсистемой, с помощью которой осуществляется формирование
хромосом при делении ядер.
Хромосомы являются основным механизмом клетки, где
собирается, хранится и выдаётся так называемая наследственная информация, включающая
в себя химическую запись последовательности фн. ячеек в структурах белков данной клетки.
Указанная информация хранится в находящихся в хромосомах молекулах ДНК. Таким образом,
молекулы ДНК представляют собой химическую запись структур всего разнообразия белков.
На длинной нити молекулы ДНК одна за другой следует запись информации о
последовательности фн. ячеек структур разных белков. Отрезок ДНК, содержащий информацию
о структуре одного белка, принято называть геном. Молекула ДНК представляет собой
собрание нескольких сот или тысяч генов. Диаметр хромосом невелик и составляет в среднем
140 ![]() , длина же
их, повторяя длину молекул ДНК, может быть свыше 1 мм. В середине периода интерфазы
происходит синтез ДНК, в результате которого хромосома удваивается.
, длина же
их, повторяя длину молекул ДНК, может быть свыше 1 мм. В середине периода интерфазы
происходит синтез ДНК, в результате которого хромосома удваивается.
Важнейшая функция хромосом - быть хранительницей записей
структур и, соответственно, алгоритмических способностей фн. подсистем клетки с помощью
механизма образования белковых фщ. единиц. С течением времени, по мере приращения функций
того или иного вида органических систем, изменяется и совершенствуется запись в хромосомах,
следуя требованиям законов фн. развития Материи. В прямой зависимости от молекулярной
записи ДНК хромосом через механизм синтезирования белковых молекул происходят все
процессы жизнедеятельности клеток. Число хромосом постоянно для каждого вида животных
и растений, то есть в каждой клетке любого организма, принадлежащего к одному виду,
содержится совершенно определённое число хромосом (рожь - 14, человек - 46, курица -
78 и т.д.). Хромосомный набор, содержащийся в ядре одной клетки, имеет всегда парные
хромосомы. Так, 46 хромосом человека образуют 23 пары, в каждой из которых объединены
две одинаковые хромосомы. Хромосомы разных пар отличаются друг от друга по форме и месту
расположения. В результате митоза получаются две дочерние клетки, по строению полностью
сходные с материнской. Каждая из них имеет точно такие же хромосомы и такое же их число,
как и материнская клетка. Таким путём обеспечивается полная передача всей наследственной
информации каждому из дочерних ядер. Ядро и все органоиды цитоплазмы клетки
взаимодействуют как единая система.
Все клетки имеют сходный тип строения: ядро, митохондрии,
комплекс Гольджи, эндоплазматическую сеть, рибосомы и другие органоиды. Однако, прежде
чем стать столь совершенной системой, какой она является в наши дни, клетка прошла
длительный путь эволюции, отмеченный соответствующими отрезками на ординатах t и
ft тензора Развития Материи. Вначале она была частью неизвестных нам
неклеточных организмов, затем несовершенных одно- и многоклеточных, включая бактерии
и синезелёные водоросли, и в конечном итоге достигла совершенства сложного клеточного
механизма, характерного для представителей современного нам растительного и животного
мира. Вследствие движения Материи по ординате качества в процессе эволюции клетки
образовалось огромное разнообразие её типов, каждый из которых был наделён строго
определёнными фн. свойствами и соответствует определённой точке на данной ординате.
Вместе с тем, с определённого момента этот процесс стал
протекать одновременно с началом развития фн. систем более высокого организационного
уровня, фн. ячейки которых клетки стали заполнять в качестве фщ. единиц. В итоге клетка
превратилась в сложное системное образование, для поддержания фн. свойств которого внутри
и вне её постоянно протекают сложные химические процессы. Постоянство процессов связано с
тем, что время функционирования фщ. единиц по мере увеличения их молекулярного веса всё
более не совпадает со временем существования фн. ячеек структур, которые они заполняют,
так как при ограниченном пространстве перемещения фщ. единиц время их существования
находится в прямой зависимости от их фщ. массы. Кроме того, постоянство процессов вызвано
тем, что большинство протекающих в клетке химических реакций имеет необратимый характер.
Для всех этих реакций свойственны величайшая организованность и упорядоченность: каждая
реакция протекает в строго определённом месте в строго определённое время в строго
определённой последовательности. Молекулы ферментов расположены на мембранах митохондрий
и эндоплазматической сети в том порядке, в котором идут реакции.
В клетке имеются около тысячи ферментов, с помощью которых
протекают два типа реакций: синтеза и расщепления. Основным (созидающим)
типом реакций следует считать реакции синтеза, в процессе которых образуются сложные
молекулярные соединения, заполняющие в качестве фщ. единиц фн. ячейки подсистемных
структур клетки. Так, на замену каждой отфункционировавшей молекулы белка, покинувшей
ту или иную фн. ячейку, на освободившееся место поступает новая молекула белка, по
структуре и химическому составу, а следовательно и своим фн. свойствам, полностью
адекватная предыдущей фщ. единице. Это означает, что вновь синтезированная фщ. единица
способна (или должна быть способна) принять полноценное участие в любом из характерных
для данной фн. ячейки алгоритмов.
Синтезирование фщ. единиц осуществляется с помощью
функционирования специальных подсистем клетки на основе кодированной генозаписи ДНК.
Имеющие при этом место флуктуационные отклонения в случае их положительного эффекта
обратной связью фиксируются в генозаписи и служат целям дальнейшего совершенствования
данной системной структуры. При отрицательном эффекте от вновь синтезированной фщ.
единицы нарушается выполнение части фн. алгоритмов и, в случае, если система не в
состоянии это элиминировать, ненадлежащее функционирование соответствующей подсистемы
в конечном итоге может привести к разрушению структуры данной клетки в целом. Таким
путём организационное устройство клетки позволяет ей поддерживать постоянное наличие
соответствующих фщ. единиц в фн. ячейках своих подсистем, что сохраняет её структуру
и чем обеспечивается способность клетки выполнять алгоритмы фн. ячеек систем более
высокого порядка, куда она входит в качестве фщ. макроединицы. Все реакции биосинтеза
(реакции ассимиляции) идут согласно общей теории систем с поглощением энергии движения
в пространстве, которая, как бы увязая в структуре системы клетки, преобразуется в
энергию связи между её фн. ячейками.
Другой тип реакций - реакции расщепления - происходит с
одновременным уменьшением энергии связи, преобразующейся в энергию движения. При
реакциях диссимиляции фщ. единицы подсистем клетки, являющиеся системными образованиями
более низкого порядка, отфункционировав, распадаются на фщ. единицы своего подуровня,
готовые при необходимости вступить в новые синтезирующие реакции с целью формирования
новых структур - фщ. единиц более высокого организационного уровня. Оба типа реакций
тесно взаимосвязаны и представляют собой единый процесс, направленный на заполнение
фн. ячеек структуры клетки дееспособными соответствующими фщ. единицами, что в итоге
обеспечивает поддержание на должном уровне фн. свойств клетки, как единого целого.
Одним из основных и сложнейших типов синтезирующих реакций
является биосинтез белков, протекающий в клетке постоянно в течение всего периода
времени её существования. В процессе функционирования клетки часть её белков, принявших
участие в каталитических реакциях, постепенно денатурируется, их структура, а
следовательно и функции, нарушаются и они удаляются из своих фн. ячеек, а затем и из
самой клетки. Их место в фн. ячейках занимают вновь синтезированные белковые молекулы,
по своим фн. свойствам полностью идентичные освободившим им место фщ. единицам. Учитывая,
что существует великое множество типов белковых молекул, механизм их синтезирования,
совершенствовавшийся в течение долгого времени, в конечном итоге превратился в
специализированную подсистему клетки с чётким перечнем алгоритмов функционирования.
Программа синтеза белков, то есть информация об их структуре,
записанная и хранящаяся в ДНК, направляется в рибосомы с помощью информационных РНК
(и-РНК), синтезирующихся на ДНК и точно копирующих её структуру. Каждой аминокислоте
соответствует участок цепи ДНК из трёх рядом стоящих нуклеотидов: А-Ц-А (цистеин),
Т-Т-Т (лизин), А-А-Ц (лейцин) и т.д. Число возможных комбинаций из 4-х нуклеотидов
по 3 равно 64, хотя используемых аминокислот всего 20. Последовательность нуклеотидов
и-РНК в точности повторяет последовательность нуклеотидов одной из цепей генозаписи,
при этом с каждого гена можно снять любое число копий РНК.
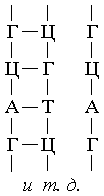 Переписывание информации на и-РНК, то есть процесс "транскрипции", происходит
при одновременном синтезировании и-РНК, которое осуществляется с помощью принципа
комплементарности. В результате образующаяся цепочка и-РНК по составу и последовательности
своих нуклеотидов представляет собой точную копию состава и последовательности нуклеотидов
одной из цепей ДНК. Молекулы и-РНК направляются затем к рибосомам, куда поступают также
и аминокислоты, доставляемые извне клетки в уже готовом виде. Аминокислоты попадают
на рибосому в сопровождении транспортных РНК (т-РНК), состоящих в среднем из 70 - 80
нуклеотидных звеньев, в 4 - 7 местах комплементарных друг другу. К одному из концов
т-РНК присоединяется аминокислота, а в верхушке изгиба находится триплет нуклеотидов,
который соответствует по коду данной аминокислоте. На каждую аминокислоту имеется своя
т-РНК, то есть их также 20 разновидностей.
Переписывание информации на и-РНК, то есть процесс "транскрипции", происходит
при одновременном синтезировании и-РНК, которое осуществляется с помощью принципа
комплементарности. В результате образующаяся цепочка и-РНК по составу и последовательности
своих нуклеотидов представляет собой точную копию состава и последовательности нуклеотидов
одной из цепей ДНК. Молекулы и-РНК направляются затем к рибосомам, куда поступают также
и аминокислоты, доставляемые извне клетки в уже готовом виде. Аминокислоты попадают
на рибосому в сопровождении транспортных РНК (т-РНК), состоящих в среднем из 70 - 80
нуклеотидных звеньев, в 4 - 7 местах комплементарных друг другу. К одному из концов
т-РНК присоединяется аминокислота, а в верхушке изгиба находится триплет нуклеотидов,
который соответствует по коду данной аминокислоте. На каждую аминокислоту имеется своя
т-РНК, то есть их также 20 разновидностей.
Синтез белков и нуклеиновых кислот происходит на основе реакций
матричного синтеза. Этим обеспечивается придание вновь образованным соединениям фн.
свойств заменяемых фщ. единиц. Новые молекулы синтезируются в точном соответствии
с планом, заложенным в структуре уже существующих молекул. Поэтому в этих реакциях
обеспечивается точная, строго специфичная последовательность мономерных звеньев
в синтезируемых полимерах. Здесь происходит направленное стягивание мономеров в
определённое место клетки - в фн. ячейки вновь образуемого полимера, при этом
расположение самих фн. ячеек предопределяется структурной организацией копируемой
матрицы. Роль матрицы в матричных реакциях играют макромолекулы нуклеиновых кислот ДНК
и РНК. Мономерные молекулы (нуклеотиды или аминокислоты) в соответствии с принципом
комплементарности располагаются и фиксируются на матрице в строго определённом, заданном
порядке. Затем происходит "сшивание" мономерных звеньев в полимерную цепь,
и готовый полимер сбрасывается с матрицы. После этого матрица готова к сборке новой
полимерной молекулы. При помощи матричного типа реакций осуществляется воспроизведение
однотипных соединений - фщ. единиц данной системы. Потребность воспроизведения однотипных
фщ. единиц прослеживается на всех уровнях организации Материи и является одной из главных
закономерностей общей теории систем.
Информация о структуре белка, записанная в и-РНК в виде
последовательности нуклеотидов, переносится далее в виде последовательности аминокислот
в синтезируемую полипептидную цепь, то есть происходит процесс "трансляции".
По мере сборки белковой молекулы рибосома ползёт по и-РНК, за ней вторая, третья и т.д.
Каждая из них синтезирует один и тот же белок, запрограммированный на данной и-РНК.
Когда рибосома пройдёт и-РНК от конца до конца - синтез белка окончен. После этого
рибосома идёт на другую и-РНК, а белок по эндоплазматической сети направляется
в соответствующую его фн. свойствам свободную фн. ячейку, которую он заполняет
в качестве фщ. единицы.
Синтез белков в клетке происходит непрерывно. Все рибосомы,
помещающиеся одновременно на одной и-РНК, объединяются в полирибосому. Рибосома работает
вдоль и-РНК "шажками": триплет за триплетом РНК находится в контакте с нею. Для
сшивания полипептидной цепи в рибосоме имеется белок - синтетаза. Молекулы т-РНК, проходя
через рибосому, задевают своим кодовым концом место контакта рибосомы с и-РНК. Если
кодовый триплет т-РНК окажется комплементарным к триплету и-РНК, аминокислота,
доставленная т-РНК, переходит из её фн. ячейки в фн. ячейку синтезируемой молекулы
белка, таким образом становясь фщ. единицей её структуры. Этим обеспечивается наиважнейшее
правило построения фн. систем - помещение данной фщ. единицы в строго соответствующую ей
фн. ячейку или, наоборот, заполнение фн. ячейки строго соответстующей ей фщ. единицей.
Поэтому имеющийся в любой клетке механизм синтезирования белков обеспечивает стопроцентную
гарантию того, что переносимая т-РНК данная аминокислота попадёт только в соответствующую
ей фн. ячейку структуры белка или, наоборот, того, что в подошедшую в рибосоме
незаполненную очередную фн. ячейку синтезируемого белка попадёт только
соответствующая ей по фн. свойствам фщ. единица - требуемая аминокислота.
Заполнив очередную фн. ячейку синтезируемого белка, рибосома
делает ещё один шаг по и-РНК, получая таким образом информацию о фн. свойствах следующей
фн. ячейки заполняемой структуры. т-РНК с освободившейся рабочей т-фн. ячейкой уходит
во внутриклеточное пространство, где захватывает соответствующую ей новую молекулу
аминокислоты с тем, чтобы вновь нести её к любой из фщ. рибосом.
Молекулы белков синтезируются в среднем около 1 - 2 мин.
Процесс этот протекает в продолжение всего периода существования клетки. Все реакции
белкового синтеза катализируются специальными ферментами, вплоть до реакций захвата
т-РНК. Все реакции синтеза эндотермичны, и поэтому каждое звено биосинтеза всегда
сопряжено с расходом АТФ.
Любая клетка сохраняет свой состав и все свои фн. свойства
на относительно постоянном уровне. Так, содержание АТФ в клетках составляет 0,04% и эта
величина стойко удерживается. Включение и выключение процессов, обеспечивающих поддержание
фн. свойств клетки, происходит в ней автоматически. Основой авторегуляции этих процессов
является специальная сигнальная подсистема клетки, использующая с этой целью фн. свойства
фщ. единиц предыдущих подуровней, то есть электромагнитные свойства электронов, атомов
и т.д. Поэтому в любой клетке всегда имеется определённое количество различных ионов и
других заряженных частиц, которые в целом создают биоэлектрические цепи, микрополя и т.п.
Изменение биоэлектрического потенциала хотя бы в одном из звеньев любой подсистемы клетки
служит сигналом для начала или окончания той или иной биохимической реакции, того или
иного перемещения фщ. единиц по фн. ячейкам различных подсистем клетки. Наличие в
структуре клеток подсистемы сигнальной биоэлектрической связи, а также использование
в этих целях фн. свойств фщ. единиц нижних подуровней (электронов, ионов и др.) служит
ещё одним подтверждением наличия тесной взаимсвязи всех уровней единой системной
организации развивающейся Материи.
Итак, конечным итогом Развития Материи по уровню З явилось
образование сложнейшей системной структуры - органической клетки. Структура каждой клетки
включает в себя строго определённое количество различных фн. подсистем, каждая из которых
выполняет строго определённую, присущую только ей функцию, обеспечивающую нормальное
функционирование всей клетки в целом. Каждая подсистема клетки имеет свою, строго
определённую структуру, включающую системные образования более низкого организационного
уровня, имеющие полимолекулярное строение со своими специфическими законами
функционирования. Каждая молекулярная структура включает в себя атомные системы со
своими специфическими законами функционирования. Атомные структуры основаны на законах
функционирования субатомных подсистем. И так бесконечно в структурную глубь Материи.
Всё указанное нагромождение фн. систем и подсистем тончайшим образом организовано в
пространстве и во времени с одной единственной целью - обеспечить проявление в строго
определённом месте в строго определённый период времени фн. свойств своеобразного
материального образования - органической клетки.
С этого момента Материя вступила в новый этап своего
качественного развития - создание саморегулирующихся и самоуправляемых макробиосистем.
![]()