Ввиду того, что развитие систем уровня З
в пространстве было ограничено пределами земной поверхности, течение
времени требовало продолжения ускоренного движения Материи по категории
качества даже тогда, когда оно почти исчерпало себя на этом организационном
уровне. Поэтому на определённом этапе Развития Материи выполнению требований этого
закона могло отвечать лишь появление новых структурных формирований, составленных
из групп клеток и обладающих иным спектором фн. свойств. Таким образом, с появлением
клетки, то есть с того момента, как она приобрела первоначальную системную законченность,
Развитие Материи по ординате качества стало переходить в следующий организационный
уровень - И, в котором уже сами клетки стали служить в качестве фщ. единиц,
заполняя фн. ячейки более сложных структур нового уровня. Это выразилось прежде всего
в фн. специализации отдельных подсистем клетки, которая с течением времени привела к
появлению многочисленных типов клеток, каждый из которых обладал строго определёнными
фн. свойствами. Поэтому движением Материи по ординате качества в пределах организационного
уровня И следует считать функциональную дифференциацию клеток, автоматически
ведущую, в силу действия 1-го принципа построения систем, к их структурной интеграции.
Следует также отметить, что согласно законам Развития Материи
количественное прибавление однотипных фщ. единиц с одинаковыми фн. свойствами не может
обеспечить заполнения вновь образующихся в процессе движения Материи по ординате
качества фн. ячеек, а значит и Развития Материи в целом. Лишь появление фщ.
единиц с различными фн. свойствами удовлетворяет этим требованиям. Однако всё разнотипное
требует обязательной системной организации. Вот почему по мере Развития Материи происходит
создание на базе имеющихся всё новых фщ. единиц с отличными от уже существующих фн.
свойствами, для реализации которых формируются структурные образования всё более
высокого системного уровня.
Именно это привело в конечном итоге к необходимости зарождения
нового вида структур, включающих клетки в свои фн. ячейки в качестве фщ. единиц. На
ординате времени этот момент был отмечен 2 - 3 млрд. лет тому назад, когда
зафиксировано, по имеющимся данным, появление Жизни на Земле. До этого
Земля, как считается, была стерильна. Однако по канонам современной биологии любое
живое существо рождается только от своих родителей, то есть от живых же существ. Поэтому
теория системного Развития Материи помогает однозначно ответить и на этот вопрос.
Вступление Материи в своём Развитии на новый этап сопровождалось
появлением многочисленной разновидности организмов растительно-животного мира. Следуя
принципам системного построения, клетки, заполняя фн. ячейки всё новых структур и
функционируя в них в качестве фщ. единиц, создавали различного рода системные и
подсистемные формирования, фн. нагрузкой многих из которых было лишь поддержание
в организованном состоянии системы клеток в процессе их специализации для образования
в дальнейшем более совершенных организмов. Эволюция растительного и животного мира
происходила в течение сравнительно длительного периода времени и основные её этапы
хорошо известны. Вместе с тем, на всём протяжении этого развития от водорослей и
бактерий до современных нам представителей флоры и фауны все процессы их образования,
существования и отмирания подчинялись единым принципам системной организации Материи,
действие которых распространяется на каждый организационный уровень, включая и подуровень
И. Все относящиеся к нему организмы представляют собой целостные системы, структуры
которых можно представить в виде определённым образом расположенных в пространстве
фн. ячеек, заполненных в качестве фщ. единиц органическими клетками.
Системы организмов имеют, как правило, фн. подситемы - органы,
несущие ту или иную фн. нагрузку. Структуру органов составляют фн. ячейки с примерно
однотипными фн. алгоритмами и поэтому заполняющие их фщ. единицы - клетки имеют
приблизительно одинаковое строение и, соответственно, фн. свойства. Группы таких клеток
носят название тканей. Как и в предыдущих организационных подуровнях, время
существования фн. ячеек не совпадает с периодом функционирования фщ. единиц. Поэтому
все организмы имеют подсистемы, обеспечивающие доставку комплектующих элементов -
различных атомов и молекул для образования новых фщ. единиц, идентичных заменяемым
в фн. ячейках отфункционировавшим фщ. единицам. При этом фн. свойства вновь образованных
клеток должны полностью совпадать с фн. свойствами заменяемых и, в конечном счёте,
корреспондировать алгоритмам заполняемых фн. ячеек. Механизмом, обеспечивающим
поддержание в фн. ячейках подсистем организмов в постоянной фн. готовности
соответствующих фщ. единиц, является митоз (деление) клеток.
Известно, что в любом организме, как и в любой фн. системе,
каждую фн. ячейку занимает строго соответствующая ей по своим фн. свойствам фщ. единица.
И наоборот, каждая фщ. единица должна занять место в строго соответствующей ей фн. ячейке.
Поэтому любое отклонение от этого правила всегда ведёт к тому, что не соответствующая
данной фн. ячейке фщ. единица не в состоянии выполнять предписания имеющихся алгоритмов
функционирования, что влечёт за собой нарушение функционирования той или иной подсистемы
организма или всей его системы в целом и что, в конечном итоге, может привести к его
гибели.
Зарождение так называемой живой природы произошло
в водах мирового океана, или вернее, на стыке морей и суши. Наличие всех компонентов,
включая воду, а также атомов большинства химических элементов в совокупности с
каждодневным постоянным источником энергии - лучистой энергией Солнца - создало идеальные
предпосылки для системного конструирования различных структур фн. ячеек, которые тут же
могли заполняться необходимыми фщ. единицами. И поэтому не эпизодические грозовые разряды
(бывшие лишь необходимым условием, но отнюдь не причиной) послужили толчком к зарождению
сложных биоструктур (как утверждают отдельные научные гипотезы), а последовательное
перебирание различных системных вариантов в сочетании с соответствующими благоприятными
условиями внесистемной среды привело к созданию динамически устойчивых биосистем. Молекулы
морской воды в сочетании с различными химическими элементами в виде растворов проникали
сквозь оболочку новых системных формирований и заполняли в качестве фщ. единиц
соответствующие фн. ячейки их структур, при этом лучистая энергия Солнца, преобразуясь
и застывая в виде энергии межмолекулярной связи, способствовала удержанию на период
функционирования фщ. единиц в своих фн. ячейках.
В результате длительного организационного процесса, протекавшего
многие миллионы лет, вначале появились простейшие одноклеточные организмы - синезелёные
водоросли и бактерии, затем зелёные водоросли, грибы и другие многоклеточные растения,
имевшие самое примитивное строение, но являвшиеся венцом творения Материи на тот момент
её Развития. Последующее течение времени и соответствующее продвижение Материи
по ординате качества требовало дальнейшего умножения функций
![]() ).
).
Мы не будем подробно описывать весь длительный процесс эволюции
организмов и их фн. подсистем в тот долгий период. Для нас важно отметить, что в
результате этого процесса появилось большое количество разнообразных растений, которые
мы отнесём к одной группе так называемых организмов первого поколения. Несмотря
на кажущееся внешнее различие, а также наличие несхожих фн. подсистем, всех их объединяет,
и это особенно важно, единый принцип построения фщ. структур. А именно: в их фн. ячейки
в качестве фщ. единиц поступают в виде растворов представители всего набора подуровней
В и Г - атомы, молекулы, ионы, радикалы и т.п., то есть элементы
неорганических соединений, находящихся в почве, а точнее, в окружающей среде и соединяемых
в фн. ячейках данного вида организмов с помощью энергии Солнца в системы очень сложной
организации. Синтезированные таким образом из CO2, H2O и других
системных образований нижних подуровней глюкоза, аминокислоты, а затем углеводы, белки,
нуклеотиды и т.д., то есть фщ. формирования более высоких подуровней заполняли в качестве
фщ. единиц фн. ячейки подсистем органических клеток, которые сами уже являлись фщ.
единицами в структуре организмов растений. Клетки, системная организация которых позволяла
производить синтез структур указанным образом, впоследствии стали называться автотрофными.
Характерными их представителями являются клетки современных нам зелёных растений.
Основной реакцией, протекающей в организмах первого поколения,
является реакция фотосинтеза:
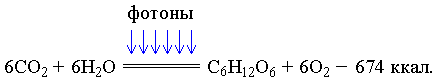
Кванты света, бомбардируя молекулярную структуру хлорофилла, передают определённое
количество своей кинетической энергии части его электронов, переводя их таким образом
в возбуждённое состояние. В результате этого электроны покидают свои орбитали
и перескакивают в более высокие. Часть из них, присоединяясь к ионам водорода, превращает
их в водород и т.д. Одновременно при этом процессе АДФ превращается в АТФ, а CO2
в глюкозу. Фотосинтез служит основой постоянно протекающего в природе великого созидательного
процесса биосинтеза, в результате которого создаётся бесчисленное множество фщ. единиц,
заполняющих соответствующие им фн. ячейки в структурах различных биоорганизмов.
В настоящее время с помощью фотосинтеза на Земле ежегодно
связывается в более сложные структуры свыше 170 млрд. тонн углерода, миллиарды тонн азота,
фосфора, серы, кальция, магния, калия и других элементов. В результате этого образуется
около 400 млрд. тонн различного рода органических веществ. Все они в виде фщ. единиц
заполняют фн. ячейки клеток всех организмов растительно-животного мира, обеспечивая их
нормальное функционирование в качестве системных образований более высокого порядка.
В процессе эволюции организмов первого поколения происходило
всё большее обособление отдельных подсистем их структур. Особенно это стало необходимым
после постепенного освоения растениями суши и приспособления к новым для себя условиям
существования. В итоге этого длительного процесса фн. дифференциации в структуре
организмов растений появились следующие органы (или подсистемы): корни, стебли, листья
и т.д., каждый из которых имеет своё фн. назначение. Так, основная функция подсистемы
корней - обеспечивать снабжение всей системной структуры организма растения фщ.
единицами предыдущих подуровней. Через корневую систему в растения поступают в виде
растворов молекулы воды совместно с атомами и ионами различных неорганических веществ,
необходимых при синтезировании сложных органических формирований (клеток, тканей и т.п.).
Поэтому фн. алгоритмы подсистемы корней должны обеспечивать постоянное устойчивое
поступление требуемых химических элементов, осуществляя при этом их опознование,
дозирование, отсортировку и транспортировку в отведённые для них фн. ячейки структуры
организмов.
По мере совершенствования корневой подсистемы в некоторых
организмах её структура стала включать также фн. ячейки аккумулятивного центра, в
которых временно размещался запас необходимых для организма растения химических элементов
и соединений. Поэтому в периоды отсутствия по какой-либо причине поступлений необходимых
элементов извне, растение могло пополнять их из аккумулятивных клеток корнеплодов. Фн.
подсистема корней является неотъемлемой частью единой структуры организма растения и
подчиняется его внутреннему алгоритмическому распорядку, направленному на обеспечение
фн. свойств растения, как целостной системы - фщ. единицы более высокого уровня. Если
произвести искусственное отделение подсистемы корней от остальных подсистем организма
растения, то внутренний алгоритмический порядок нарушится и обе части системы прекратят
своё фн. существование, распавшись на составляющие их фщ. единицы.
Другой важной подсистемой организмов растений являются
листья. Их основная функция состоит в осуществлении важнейшего органического
процесса - реакции фотосинтеза в период функционирования организмов растений. Структура
каждого листа (то есть пространственное расположение его фн. ячеек) представляет собой
довольно совершенный механизм, позволяющий обеспечивать оптимальное течение реакций
фотосинтеза при данных условиях. При этом все другие подсистемы организма способствуют
нормальному протеканию этого процесса. Полученные в результате фотосинтеза органические
соединения транспортируются в отведённые для них соответствующие фн. ячейки, освобождая
место для образования новых единиц органических соединений. Реакция фотосинтеза
сопровождается интенсивным газообменом, для чего в структуре листьев имеются
специализированные фн. ячейки с соответствующими алгоритмами, в которых происходит приток
молекул углекислого газа и отток молекул кислорода. Кроме того, в функции подсистемы
листьев входит терморегулирование реакции фотосинтеза, достигаемое путём сбора всей
излишне поступающей энергии фотонов Солнца и отвода её специальным механизмом подсистемы,
в основе действия которого лежит принцип испускания (испарения) молекул воды.
Подсистема листьев, следуя климатическим колебаниям,
функционирует лишь в благоприятные для этого периоды. Когда же температурный режим
окружающей среды препятствует нормальному течению фотосинтеза и действует разрушающим
образом на тонкие механизмы листьев, внутренний алгоритмический распорядок организма
растения предусматривает их отторжение. Это самозащитное явление нисколько не нарушает
целостного единства структуры организма растения и служит целям обеспечения сохранности
остальных его подсистем. Поэтому опадание листьев является таким же естественным событием
в цикле алгоритмов развития растений, как и их появление в процессе регенерации.
Следующей функционально важной подсистемой организмов растений
являются стебли. Перечень функций, выполняемый сочетаниями их фн. ячеек, также
весьма обширен. Сюда прежде всего следует отнести внутрисистемные пространственные
перемещения различных фщ. единиц из одних частей системы в другие: от листьев к корням,
от корней к листьям и т.д. Для этих целей структура стеблей предусматривает наличие
специальных транспортных артерий, или сосудов, пронизывающих подсистемы всего организма,
и по которым фщ. единицы перемещаются из одних фн. ячеек в другие. При этом вода и
минеральные соли поднимаются от корней в верхнюю часть растений по внутренним сосудам,
а образовавшиеся в листьях органические вещества транспортируются по внешним артериям
стеблей. У многих растений структура стеблей (стволов) включает аккумулятивные фн. ячейки,
куда складируется запас необходимых для последующего использования элементов. Стебли
(стволы) растений служат также целям оптимального расположения фн. ячеек структуры
организма растения в геометрии пространства. Поэтому даже пространственное расположение
лиственного покрова растения с целью обеспечения максимальной площади его облучения
Солнцем входит в функцию стеблей.
Ещё одной очень важной особеностью строения стеблей является
включение в их структуру сигнальной подсистемы организма растения, имеющей
свои ответвления практически во всех его органах. Однако главные каналы связи проходят
именно через стебли. По этим каналам внутренняя информация организма поступает из одной
подсистемы в другую, координируя таким образом во времени начало и прекращение тех или
иных реакций, запрограммированных алгоритмами соответствующих фн. ячеек. Эти же сигналы
служат для внесения коррекции в указанные алгоритмы. Здесь следует отметить, что само
понятие организм включает в себя наличие условно целостной
биологической системы с обязательным присутствием сигнальной подсистемы. Именно благодаря
сигнальной подсистеме некое скопление органических клеток объединено в систему единого
организма. В простейших организмах растений сигнальная подсистема появилась вначале также
в довольно зачаточном состоянии, развившись со временем в примитивную первую сигнальную
подсистему, положившую одновременно начало появлению духовности в
организме. Как уже отмечалось, сигнальная подсистема организмов растительно-животного
мира имеет биоэлектрическую природу. С её помощью происходит тесное взаимодействие
подсистем единой структуры организма, регулирование во времени алгоритмической
деятельности тех или иных фщ. единиц.
Здесь необходимо также отметить и то, что в столь сложных
системных образованиях, каковыми являются организмы I-го поколения, своё дальнейшее
развитие получило общее для всей живой организации Материи свойство - раздражимость.
Под раздражимостью понимают способность системы отвечать на внешние воздействия такой
реакцией, которая по своей силе, месту и характеру не соответствует силе, месту и
характеру самого внешнего воздействия, при этом данная реакция имеет обратимый характер,
что способствует её многократному повторению. В организмах, даже самых примитивных,
раздражимость проявляется в гораздо более сложной, чем в изолированном белковом комплексе,
дифференцированной форме, имеющей своё определённое функциональное значение, однако и
здесь она базируется на закономерностях, свойственных всем системным формированиям, а
именно: перемещении в определённый период времени отдельных фщ. единиц из одних фн. ячеек
в другие. Элементарной формой раздражимости является способность находящегося в клетках
миозина отвечать сокращением при воздействии на него минимальным количеством АТФ, как
естественного химического раздражителя. Реакция сократительного белка на АТФ исчезает,
если блокировать одну из важнейших реактивных групп белков - сульфгидрильную группу.
Восстановление этих групп в структуре сократительного белка восстанавливает и реакцию
белка на названный раздражитель. Растения не имеют специальных тканей или какого-либо
координационного центра, воспринимающих и проводящих раздражения. Однако, несмотря на
относительную примитивность реакций растений на раздражения, сложнейшая подсистема
плазматических, сосудистых и гормональных связей, объединённая в примитивную сигнальную
подсистему, в свою очередь объединяет все их части и органы в единый целостный организм
и регулирует все физиологические и биологические процессы. Возбуждённый участок ткани или
органа растения приобретает отрицательный по отношению к невозбуждённым участкам заряд,
вследствие чего между возбуждённым и невозбуждённым участками возникает электрический
ток (биоэлектрический потенциал). Кроме того, в возбуждённом участке образуются (или
освобождаются) вещества высокой физиологической активности (ауксины и другие фитогормоны),
которые передвигаются к другим участкам ткани и, наряду с биотоками, вызывают в них
состояние возбуждения. Скорость распространения возбуждения у растений составляет единицы
и десятки микронов/сек. Претерпев соответствующие молекулярно-физические изменения в ответ
на воздействие раздражающих агентов, белковые структуры в силу действия имеющейся
генозаписи их исходного построения вновь возвращаются в своё первоначальное состояние
и могут снова реагировать на те или иные воздействия. Энергия ответной реакции на
раздражение обычно пропорциональна, но не равна энергии раздражения, так как реакция
на раздражение осуществляется за счёт внутренней энергии организма растения, накопленной
ранее при ассимиляции. Если в предыдущих реакциях на раздражения эта внутренняя энергия
израсходовалась, то новые раздражения не будут вызывать ответной реакции до тех пор,
пока не восстановятся исходный энергетический уровень и другие свойства возбуждаемого
участка ткани. Очень сильные раздражения не стимулируют, а наоборот, угнетают
жизнедеятельность организма, и при достаточной продолжительности действия такие
раздражители нарушают нормальный ритм его функционирования. В силу этого сила
раздражения должна быть строго дозирована.
Организмы I-го поколения, несмотря на их относительную
примитивность, имели уже довольно надёжную подсистему алгоритмозаписи, в основе
которой лежит биохимическая запись генетического кодирования ДНК. В ней собирается
информация практически от всех клеток, входящих в организм. По мере усложнения системной
организации растений повышалась надёжность и подсистемы алгоритмозаписи, обеспечивавшей
кодирование развёртывания структуры фн. ячеек всех подсистем организма, соотнесённое
с пространственно-временными интервалами. Вначале подсистему алгоритмозаписи имел
практически каждый орган растения. Так до сих пор существуют растения, у которых при
культивировании лишь одного органа происходит развёртывание всех остальных. К ним можно
отнести лесной ландыш (корневище), тополь (часть стебля) и т.д. Однако, в конечном итоге,
наиболее надёжной оказалась система записи алгоритмов, производимая в особом, специально
для этого предназначенном органе растения - его семенах. Одно из главных
преимуществ такой записи является возможность её реализации (алгоритмочтения) с
большим интервалом как в пространстве, так и во времени.
И действительно, семена можно перенести на многие километры
от материнского растения и посадить, то есть дать начало развития нового организма
растений, через несколько лет после отделения семени от родителя. Всё это отвечало
требованиям Развития Материи по ординатам качества-времени-пространства. Мы
не будем останавливаться на самом механизме алгоритмической записи развёртывания
структур подсистем всего организма растения в зародыше семян, однако отметим, что запись
эта настолько полная, что включает в себя количественное и качественное отличие всех
входящих в структуру данного организма фн. ячеек, время их развёртывания и период
функционирования, а также алгоритмические особенности каждой группы функционально
обособленных ячеек. Поэтому как только семя попадает в соответствующую фн. ячейку
биогеоценоза, тут же включаются его биочасы и начинается декодирование кропотливо
составленной генозаписи зародыша, являющееся первой фазой развёртывания структуры
организма очередного растения.
Семена, как известно, помимо генетической записи зародыша
имеют и небольшой запас (сухой паёк) тщательно отобранных элементов, необходимых для
использования в качестве фщ. единиц на первых порах развёртывания структуры растения.
Позднее, по мере развития их различных подсистем, организмы растений стали более
запасливыми и помимо накопления строго обязательного запаса необходимых
элементов в семени, они начали также аккумулировать значительное количество элементов
в другой своей, более обширной аккумулятивной подсистеме - плодах. При созревании
плодов основная масса их фн. ячеек, имеющих главным образом аккумулятивную функцию,
заполняется всеми элементами, необходимыми для нормального развёртывания из семян первых
подсистем растения. Это заполнение, как и все трансформации в растениях, происходит не
хаотически, а повинуясь строгому регламентированию соответствующими алгоритмами, согласно
которым строго определённые молекулярные соединения в виде фщ. единиц заполняют отведённые
для них фн. ячейки, где они с помощью энергии Солнца полимеризуются в более сложные
соединения, что обеспечивает им более продолжительный период существования. В последующем,
после завершения созревания плода и семян, то есть когда все фн. ячейки их структур
наполнятся соответствующими фщ. единицами, плод вместе с семенами опадает на верхний
слой почвы, где происходит деполимеризация его фщ. единиц, в результате чего создаётся
среда питательных элементов для находящихся здесь же семян. Поэтому, как только из
семени начинается развёртывание структуры нового растения, сохранившиеся элементы
деполимеризированного плода служат основным источником, обеспечивающим заполнение
его фн. ячеек соответствующими фщ. единицами.
В процессе своего формирования каждое семя проходит стадию
оплодотворения, то есть момент соединения двух системообразующих структур - пыльцы
и яйцеклетки. Это соединение служит целям улучшения генотипа растений посредством
распространения более совершенных структур фн. ячеек подсистем, образовавшихся при
мутации генов. Совершенствование этого процесса шло по пути от обоеполых растений через
однодомные, то есть у которых есть и тычиночные, и пестичные цветки, до двудомных, где
тычиночные и пестичные цветки расположены на разных растениях. Таким образом, уже у
организмов I-го поколения сформировались особи разных полов. Появление семян от разнополых
растений обеспечивает наличие генозаписи как минимум от двух родительских системных
образований, что способствует постоянному совершенствованию структуры фн. ячеек данного
вида растения и соответственной оптимализации совокупности их алгоритмов. Генозаписью
алгоритмов построения и функционирования фщ. единиц всех подсистем растения, произведённой
в ДНК клеток зародыша семян, а также обеспечением минимального запаса необходимых при
развёртывании структуры организма элементов, сосредоточенного в плодах, практически
заканчивается фн. деятельность большинства растений - организмов I-го поколения.
Отфункционировав, структуры их подсистем распадаются, а фщ. единицы, заполнявшие ранее
их фн. ячейки, деполимеризуясь, покрывают верхний слой почвы, образуя и поддерживая таким
образом её гумусовый слой. В дальнейшем разрозненные элементы гумусового слоя могут войти
в состав фщ. единиц структуры нового растения с тем, чтобы, отфункционировав там, вновь
вернуться в гумусовый слой. Этот процесс бесконечен и составляет основу биогеоценоза.
Как ни велико множество разновидностей организмов I-го
поколения, функциональная нагрузка их в целом одинакова, а разница состоит лишь
в структурной организации их подсистем, приспособленных под те или иные особенности
биогеоценоза, в котором они территориально размещены и фщ. единицами которого они
сами являются. Поэтому, исчерпав весь набор возможных функциональных приращений
![]() )
)
К II-му поколению организмов относятся все растительноядные
представители животного мира. Развитие у них подсистемы ускоренного искусственного
расщепления органических соединений тканевых структур растений позволило им добывать
в большом количестве комплексные материальные соединения, с помощью которых они могли
постоянно заполнять фн. ячейки своих всё более усложнявшихся подсистем, что способствовало
появлению фн. ячеек с новыми свойствами и соответствовало движению Материи по ординатам
качества-времени. Мы не будем подробно анализировать эволюцию организмов II-го
поколения от простейших одноклеточных до современных нам хордовых класса млекопитающих
травоядных животных. Отметим лишь, что главной причиной дивергенции их системной
организации была потребность следования законам Развития Материи. Основой этого
длительного процесса явилось усложнение морфофизиологической структуры организмов,
что привело к появлению в протерозойскую эру (2 млрд. лет назад) животных с двусторонней
симметрией тела с дифференцировкой его на передний конец и задний. Передний конец стал
местом для развития органов чувств, нервных узлов, а в дальнейшем - головного мозга.
В процессе последующей эволюции происходила, главным образом, дивергенция типов
животного мира и замена первоначальных низкоорганизованных примитивных форм более
высокоорганизованными путём ещё большей дифференциации строения и функций тканей и
органов организмов. При этом фн. ячейки тканей организмов II-го поколения заполнялись
в качестве фщ. единиц уже лишь гетеротрофными клетками, то есть неспособными к синтезу
органических соединений из неорганических. В самих клетках ещё более усовершенствовалась
система генозаписи в цепочках ДНК. Характерной особенностью клеток любого органа осталось
то, что в каждой из них имелся весь набор генов данного вида организмов, однако в клетках
разных тканей используются лишь немногие группы генов, то есть только те из них, на
которых записаны алгоритмы структурного развёртывания и функционирования структур фн.
ячеек, которые данные клетки занимают в качестве фщ. единиц.
Протекавший многие сотни миллионов лет морфофизиологический
прогресс, или ароморфоз, привёл к значительным эволюционным изменениям подсистем структуры
организмов II-го поколения (что выразилось в общем подъёме их организации), биологическому
прогрессу, а также другим не менее важным последствиям. Сюда прежде всего следует отнести
отрыв их систем от гумусового слоя почвы и способность свободно и автономно перемещаться
по субстрату. Вследствие этого организмы получили возможность постепенно осваивать до
этого пустынные районы земной поверхности в трёх сферах: на суше, в воде и воздухе, что
вело к увеличению фн. разнообразия их структур и вполне отвечало требованиям движения
Материи в качестве-времени-пространстве. Приобретённая способность к передвижению
в пространстве околоземной поверхности позволило организмам II-го поколения перемещаться
от одного источника питания (системы организмов I-го поколения) к другому, максимально
расширяя ареал своего обитания. Кроме того, в неблагоприятные моменты организм стал иметь
возможность укрываться в более безопасном для себя месте. Потребление разнообразных
травянистых растений увеличило набор элементов, из которых слагались фщ. единицы,
заполнявшие фн. ячейки подсистем организмов животных, при этом каждый элемент заполнял
отведённую именно для него фн. ячейку, где он мог проявить свои, присущие лишь ему фн.
свойства. Вместе с тем, как и во всех системных образованиях предыдущих подуровней, любая
вновь означившаяся фн. ячейка структуры того или иного организма безусловно требовала для
своего заполнения лишь фщ. единицу, способную выполнять её набор фн. алгоритмов. Малейшее
несоответстие фщ. единицы заполняемой фн. ячейке вело к нарушению функционирования данной
подсистемы организма и к возможному прекращению существования всей его системы в целом.
Рассмотрим вкратце строение организмов II-го поколения. В
качестве примера возьмём структуру организма любого современного нам млекопитающего
животного. Его целостная полуавтономная система включает в себя множество подсистем.
Одной из основных среди них является опорно-двигательная подсистема. Она включает
в себя костный скелет с прикреплёнными к нему группами мышц. Костный скелет, определяя
геометрическое расположение в пространстве прочих подсистем организма, в отдельных случаях
выполняет и защитную функцию. Клетки мышечной ткани с помощью биохимических реакций при
участии АТФ, как универсального источника биоэнергии, сокращаясь в заданный момент времени,
приводят к пространственному перемещению с заданной скоростью отдельных частей организма.
Опорно-двигательная подсистема, хорошо скоординированная и чётко управляемая, позволяет
некоторым современным животным перемещаться со скоростью в несколько десятков км/час.
Другой важной подсистемой организма является подсистема
пищеварения. Она включает в себя ряд органов, где регулярно протекают процессы
размельчения органических соединений подсистемных образований организмов I-го поколения
до такого состояния, когда они могут быть использованы в качестве составных элементов в
синтезируемых гетеротрофных клетках различных органов подсистем рассматриваемого нами
организма. Регулярность указаных процессов определяется потребостью отдельных подсистем
в замене в их фн. ячейках отфункционировавших фщ. единиц на новые. Наряду с подсистемой
пищеварения функционирует также подсистема выделения. Через её органы из организма
удаляются непотребовавшиеся элементы, имевшиеся в органических соединениях пищи, а также
элементы распада отфункционировавших фщ. единиц большинства подсистем организма.
Постоянно функционирующая подсистема дыхания
служит для обеспечения газообмена протекающих в различных органах и тканях
биохимических реакций. В процессе газообмена происходит постоянный подвод требующегося
для окислительно-восстановительных реакций кислорода, а также отвод одного из продуктов
распада всех органических соединений - углекислого газа.
Аккумулятивная подсистема организма включает в себя
органы, фн. ячейки которых заполняются определённым запасом большинства элементов,
необходимых для построения фщ. клеток других подсистем, удлиняя тем самым период
автономного функционирования организма в целом. В органах данной подсистемы скапливается
также ряд органических соединений, последующее расщепление которых может послужить
дополнительным источником энергии. Аккумулятивная подсистема имеет очень важное значение
в жизнедеятельности организмов животного мира. С её помощью организм имеет возможность
увеличить интервалы между приёмами пищи, нормально функционируя в указанные перерывы.
Это особенно важно для животных, ареал обитания которых находится в пустынной местности,
а также в зимнее время года.
Подсистема крово- и лимфообращения обеспечивает
постоянную надёжную транспортировку всех необходимых для протекающих в клетках
биохимических реакций компонентов и отвод элементов, образовавшихся в процессе распада
отфункционировавших единиц. Кровь представляет собой имеющую свойства жидкости структуру
фн. ячеек, заполненных соответствующими фщ. единицами. Поэтому в крови всегда содержится
полный перечень потребляемых в клетках при их синтезировании элементов, и они в необходимый
момент перемещаются из фн. ячеек крови в соответствующие фн. ячейки синтезируемой клетки.
Освободившиеся фн. ячейки крови тут же заполняются новыми фщ. единицами из аккумулятивной
подсистемы фн. ячеек или же непосредственно из подсистемы пищеварения. Удержание
в фн. ячейках крови соответствующих элементов и соединений, а также их перемещение
в фн. ячейки синтезируемых клеток имеет биоэлектрическую основу.
Ввиду того, что все биохимические реакции в клетках
протекают при строго заданной температуре, в организмах II-го поколения (у теплокровных
животных - млекопитающих) существует более совершенная, чем у I-го поколения, подсистема
терморегулирования, обеспечивающая постоянство внутренней температуры тела,
несмотря на любые температурные колебания внешней среды. Порой эти колебания
достигают 70oC.
Из-за большой сложности построения и функционирования системы
организмов II-го поколения, ей потребовалась надёжная подсистема самосохранения,
или защитная подсистема, зачатки которой мы можем наблюдать уже у организмов I-го
поколения. Указанная подсистема включает специальные органы и фн. алгоритмы как внешней,
так и внутренней самозащиты. В частности, внутренняя самозащита направлена, в основном,
против проникающих в различные органы организмов чужеродных образований, которые
подсистема самозащиты старается разрушить и вывести за пределы системы. Интересен один
из способов внутренней самозащиты, основанный на принципе постоянства температуры для
протекающих в биосистемах реакций. Исходя из того, что вторгнувшиеся микроорганизмы
(например, вирусы) реакционно более активны, поскольку практически не имеют аккумулятивной
подсистемы, и их системная организация в большей части менее приспособлена к колебанию
температур из-за существующей разности фн. масс, организм в целях самозащиты через
подсистему терморегулирования повышает во всей своей системе общую температуру,
сознательно идя на риск временного нарушения своих отдельных биореакций. Однако
вызванные этим нарушения в инородных микросистемах значительно серьёзнее, вследствие
чего они погибают или резко снижают свою активность, после чего и выводятся за пределы
системы организма, в то время как подсистемой терморегулирования вновь восстанавливается
характерный для данного организма температурный режим.
Организмам II-го поколения, как известно, приходится постоянно
перемещаться в поисках пищи по суше, воде или воздуху. Для обеспечения безопасного
передвижения, а также более продуктивных поисков пищи в системах этих организмов получила
широкое развитие подсистема восприятия, поиска и ориентации. Она
включает в себя органы зрения, слуха и обоняния. С их помощью организмы легко
ориентируются в пространстве и более эффективно ведут поиск потребляемых частей
организмов I-го поколения. Указанные органы участвуют также в алгоритах
функционирования подсистемы внешней самозащиты.
Среди прочих подсистем организмов II-го поколения следует
выделить три наиболее важных. Одной из них стала выделившаяся подсистема передачи
раздражимости, или возбуждения. Для перемещающегося по субстрату организма в условиях
быстро меняющейся ситуации потребовалась более ускоренная передача соответствующих
сигналов от одного органа другому. Вследствие этого передача сигналов в организмах II-го
поколения стала носить целиком биоэлектрическую основу, а выделившаяся подсистема передачи
развилась в центральную нервную подсистему (ЦНП). Входящие в эту подсистему клетки
отличаются особенно хорошей электропроводимостью, в силу чего в них постоянно циркулируют
так называемые токи покоя и токи действия. При наличии какого-либо раздражителя происходит
возбуждение данного участка ткани, в связи с чем возникает ток действия. Возбуждённый
участок ткани приобретает отрицательный электрический заряд по отношению к любому
невозбуждённому её участку, после чего биоэлектрический потенциал передаётся согласно
имеющемуся алгоритму в соответствующий орган системы, при этом скорость передачи сигнала
благодаря эволюции постепенно возросла до 120 м/сек. Единая ЦНП организмов II-го поколения
приняла на себя функцию координирования фн. деятельности практически всех подсистем
организма, являясь таким образом основой более усовершенствованной, чем у организмов
I-го поколения первой сигнальной подсистемы, а вместе с ней и своеобразной
духовности организмов. Дальнейшая эволюция первой сигнальной подсистемы
организма протекала по пути установления и закрепления так называемых рефлекторных дуг,
которые составляли определённую цепочку фн. ячеек, заполненных соответствующими нервными
клетками. В процессе формирования ЦНП её отдельные части всё более функционально
дифференцировались, образуя спинной мозг, головной мозг, вегетативную нервную подсистему.
Отличительной чертой нервных клеток является то, что они, в отличие от других, практически
не имеют способности к делению и существуют в течение всей жизни организма, в силу чего
установившаяся один раз рефлекторная дуга при определённых условиях существует до момента
распада всей системы организма. Первая сигнальная подсистема включает в себя рефлекторные
дуги, передающие возбуждения как от рецепторов, реагирующих на внешние раздражители, так
и от рецепторов внутренних раздражений. Структура устойчивых рефлекторных дуг генетически
записывается и воспроизводится в последующих поколениях, образуя перечень так называемых
безусловных рефлексов. В итоге нервная подсистема организма приобрела наибольшее значение
в осуществлении регулирования и чёткой координации фн. деятельности различных подсистем
целостного организма.
В процессе существования организмов II-го поколения стало
складываться всё больше ситуаций, при которых на раздражение отдельных рецепторов
организму целесообразнее было реагировать совершенно по разному. Так, например, сытое
животное при виде новых порций пищи или воды никак не реагирует на них, поскольку его
первая сигнальная подсистема наряду с получением сигнала от рецептора глаза одновременно
получает также сигнал и от рецептора аккумулятивной подсистемы организма и этот сигнал
по своей раздражающей силе на какое-то время оказывается сильнее первого. Для анализа
постоянно поступающих в неё сигналов о различной силы раздражениях многочисленных
рецепторов на стыках центров преломления рефлекторных дуг в недрах ЦНП стали формироваться
так называемые центры анализа и обработки раздражающих сигналов, на
которые легла функция координации хода последующих реакций на большинство раздражений,
передаваемых от различных рецепторов. По мере эволюции организмов II-го поколения эти
аналитические центры первой сигнальной подсистемы всё более локализовывались в структурах
головного мозга, а учитывая, что функционально организмы II-го поколения постепенно всё
более разнились между собой, аналогичную всё большую разницу приобретали и аналитические
фн. центры ЦНП. Таким образом, со временем становилось всё более очевидным, что каждая
вновь появляющаяся функция организмов II-го поколения получала свой, обслуживающий только
её аналитический центр головного мозга ЦНП, то есть актуальная область движения Материи
в качестве-времени ![]() )
)
Ещё одной важной подсистемой организмов II-го поколения
стала подсистема генозаписи, которая помимо кодирования структурного развёртывания
всей системы организма, а также состава всех его фщ. единиц стала генетически записывать
ещё и рефлекторные связи дуг, и соотвествующие аналитические фн. центры сигнальной
подсистемы ЦНП. Именно таким путём начал складываться генотип организмов.
Вновь образовывавшиеся впоследствии рефлекторные дуги и аналитические фн. центры
при закреплении их в качестве условных рефлексов составляли фенотип организма,
после чего генетически записывались и передавались по наследству, входя уже наряду с
ранее записанными рефлексами в генотип последующих поколений, соответственно пополняя
его и всё более развивая его духовность .
Последней важной подсистемой организмов II-го поколения
следует считать подсистему воспроизведения потомства, основанную на функциональном
разделении всех организмов на два пола: мужские и женские особи. Каждый пол со временем
приобретал всё большую фн. специализацию, однако наибольшего отличия достигли органы
подсистем, принимающие непосредственное участие в производстве потомства. Зарождение
каждого организма начинается с момента соединения двух специализированных клеток - гамет,
отдельно заимствованных от особей обоих полов. В каждой гамете имеется своя генетическая
запись, заключенная в гаплоидном наборе нескольких десятков хромосом, при этом любое
внутрихромосомное отклонение генома определённым образом отражается на формирующемся
генофонде потомства. Развитие зародышей организмов млекопитающих первое время протекает
в специальной подсистеме материнского организма под контролем его ЦНП, регулирующей
прежде всего подвод соответствующих питательных элементов для заполнения фн. ячеек
развёртываемой структуры нового организма. После рождения детёныша и отделения его от
материнской системы, снабжение нового организма питательными элементами материнским
организмом осуществляется ещё продолжительное время и они поступают в него в виде
специального раствора (молока), вырабатываемого соответствующей фн. подсистемой
организма женской особи. У организмов II-го поколения существуют также подсистемы
воспроизведения потомства посредством выкладки яиц, представляющих собой зародыш
в строго дозированной среде тщательно отобранных питательных элементов, которые он
полностью использует в качестве фщ. единиц для фн. ячеек развёртываемой структуры
до определённого момента своего развития.
Таким образом, морфологическая и физиологическая
дифференциация подсистем организмов II-го поколения, протекавшая многие миллионы
лет, отвечала потребностям движения Материи по ординате качества-времени
![]() ),
),
В процессе эволюции новые, высшие по своей организации группы
организмов возникали путём ароморфозов, идиоадаптаций и дегенераций. На одном из этапов
указанного процесса развития системной организации Материи появились представители
организмов третьего поколения. К ним относятся организмы, использующие в качестве
строительных полуфабрикатов при синтезировании своих фщ. единиц не неорганические
вещества гумусового слоя и не органические соединения размельчённых тканей отдельных
органов растений, а значительно более сложные органические вещества тканей организмов
II-го поколения. В результате этого, у плотоядных животных, как они впоследствии стали
называться, отпала необходимость постоянно и в больших количествах потреблять отдельные
органы разнообразных растений для того, чтобы заполнить фн. ячейки своих подсистем
соответствующими фщ. единицами. Им стало достаточно овладеть одним из организмов II-го
поколения, чтобы получить сразу в большом количестве разнообразие многих необходимых
элементов, находящихся в фн. ячейках организма растительноядного животного и из которых
они могли синтезировать фщ. единицы для подсистем своего организма. Начиная с этого
времени необходимые элементы организм стал получать готовыми блоками (блок-питание),
что вполне отвечало принципам построения материальных систем, предопределяющим
использование в качестве фщ. единиц в структурах всех последующих ступеней организации
устойчивых комплексов единиц предыдущих уровней.
В системной организации организмов III-го поколения произошло
меньше изменений по отношению к организмам II-го поколения, чем это было между поколениями
II-м и I-м. Прежде всего значительно изменилась подсистема пищеварения, приспосабливаясь
под новый вид питания, а также ещё большую фн. значимость получила нервная подсистема.
Среди организмов III-го поколения по уровню своего развития всё больше стали выделяться
наземные животные. В конечном итоге, всё дальнейшее развитие животного мира стало
сводиться, в основном, именно к последовательному усложнению у наземных организмов III-го
поколения ЦНП, повышению интенсивности и эффективности её использования, увеличению
разнообразия спектра её функций. Главным образом, это сказалось на системной организации
головного мозга, который всё больше становился специализированной подсистемой умножающихся
аналитических фн. центров, объединяющих анализаторы и инициаторы большинства процессов,
протекавших внутри организма, и некоторых - вне его.
Несмотря на большое количество видов организмов всех трёх поколений
(а их только в настоящее время насчитывается на Земле около 0,5 млн. видов растений и
1,5 млн. - животных) и их фн. разнородность, на ординате качества-времени тем
не менее всё равно наступил момент, когда всего этого разнообразия стало недостаточно
для обеспечения дальнейшего Развития Материи. Выход из этого мог быть, как и прежде,
найден лишь в ещё более сложной системной организации Материи путём создания очередного
нового организационного уровня. Первые предпосылки перехода к нему начали возникать
ещё около 30 млн. лет назад, когда в лесах палеогена и неогена появились парапитеки -
животные величиной с кошку, которые жили на деревьях и питались растениями и насекомыми.
От парапитека произошли современные гиббоны и орангутаны и ещё одна ветвь - вымершие
древние обезьяны дриопитеки, которые дали три ветви, приведшие к шимпанзе, горилле и
человеку. Чарльз Дарвин убедительно доказал, что человек представляет собой
последнее, высокоорганизованное звено в цепи развития живых существ четырёх поколений
и имеет общих далёких предков с человекообразными обезьянами.
Итак, результатом движения Материи по организационному уровню
И следует считать создание наиболее развитых организмов - организмов IV-го
поколения, к коим мы причисляем только человека, система организма которого в целом
к тому времени достигла стабильного совершенства. Будучи производной системой, вобравшей
в себя всё лучшее от организмов II-го и III-го поколений, человек получил в качестве
генетического наследства набор всех тех подсистем, которые обеспечивали его
существование и надёжное функционирование в широком диапазоне окружающей среды.
В качестве питания для заполнения фн. ячеек своих подсистем его организм всё более
приспосабливался к потреблению высокопитательных частей организмов I-го и II-го
поколений. Так, в его рационе всё большую долю стали занимать элементонасыщенные
аккумулятивные подсистемы, формируемые вокруг семян у организмов I-го поколения (плоды,
ягоды, фрукты) и различные части организмов II-го поколения. Части организмов III-го
поколения, то есть плотоядных животных, человек в пищу практически не потреблял и не
употребляет, как этого не делают и сами плотоядные животные, ввиду невозможности их
использования для заполнения фн. ячеек подсистем его организма. Однако, опережающее
развитие и специализацию в дальнейшем вплоть до наших дней в организме человека всё
более стала получать подсистема, регулирующая его высшую нервную деятельность, и в
первую очередь, структура его головного мозга.
И действительно, если у человекообразной обезьяны объём черепа
составлял 600 см3, то уже у первого человека, австралопитека, жившего 3 - 5
млн. лет назад, объём мозга стал составлять 800 см3. У питекантропа - 1 млн.
лет назад - объём черепа колебался уже в пределах 900–1100 см3. Благодаря
прямохождению у обезьяноподобных предков человека руки освободились от необходимости
поддерживать тело при передвижении и стали приобретать способность к другим
разнообразным вспомогательным движениям. В силу этого у питекантропа хотя ещё и не было
приспособленных жилищ, но он уже умел пользоваться огнём и начал использовать различные
предметы в качестве первых орудий. Помимо огромного преимущества, полученного в связи
с освобождением передних конечностей, переход к прямохождению давал гоминидным предкам
человека ещё одно эволюционное приобретение: в результате изменения положения головы и
глаз сильно возрос объём воспринимаемой ими зрительной информации, вследствие чего в
огромной степени расширились возможности в выработке адекватного конкретной ситуации
поведения.
Если сам переход австралопитеков к прямохождению не мог
осуществиться без сильного изменения фн. свойств их мозга, то совершенствование
прямохождения и возросшие в связи с этим возможности ориентации во внешней среде
так же, как и использование руки, в свою очередь повысили роль мозга как центральной
подсистемы оценки информации о внешней среде и управляющей поведением всего организма.
Параллельно с указанным процессом происходило анатомическое совершенствование руки как
органа трудовой деятельности, вначале ещё примитивной, но на последующих этапах эволюции
превратившейся постепенно в орган сложной, сознательно программируемой деятельности.
Несомненно, что имевший при этом место отбор опирался на
оптимальный геномный набор, контролирующий анатомическое строение органов. Вместе
с тем, адаптивное фн. использование всех анатомических завоеваний и их дальнейшее
эволюционное совершенствование были уже невозможны без совершенствования мозга как
центрального аппарата, управляющего новыми функциями тела, в силу чего основными
критериями дальнейшего отбора всё более становились структура и фн. свойства мозга.
Поэтому именно мозг как подсистема управления положением и функционированием тела,
деятельностью освобождённой руки, а также ориентации в конкретной жизненной ситуации
и построения программ поведения стал являться с тех пор главнейшим фактором
естественного отбора. Именно дальнейшее умножение и совершенствование его аналитических
фн. центров, отражавшие приращение функций ![]() )
)
![]()